Annals of Neurosciences, Volume 18, Issue 3 (July), 2011
Nuclear organization and distribution in the brain regions of the snake headed fish, Channa marulius
KEY WORDS
Nuclear groups
Channidae
Forebrain
Midbrain
Hindbrain
Corresponding Author:
Yashashree A. Gadhikar, PhD
Tel : + 91721-2531706
E-mail: yash_gadhikar@rediffmail.com
ABSTRACT
Background: Morphological variations have arisen due to diverse environmental conditions. Application of cytoarchitectonic criteria permits the delineation of distinct nuclear complexes from the brain region. Purpose: Very less information is available on the cytoarchitectonic pattern of the brain, of the giant snake headed murrel, Channa marulius. The murrel is much neglected in neuroanatomical terms and their study is a necessary step in tracing the evolutionary trends. Hence, in the present investigation, the brain of the snake headed fish, Channa marulius has been investigated to reveal the organization of different nuclear complexes. Methods: Different nuclear complexes were identified and studied using Cresyl violet and Haematoxyline-Eosin staining techniques from the brain region of Channa marulius. Results: Five distinct nuclear complexes namely pars medialis, pars centralis, pars lateralis dorsalis and pars lateralis ventralis respectively were observed in the area dorsalis telencephali and five nuclear groups pars ventralis, pars lateralis, pars dorsalis, pars supracommissuralis and nucleus entopeduncularis were identified in the area ventralis telencephali. Conclusions: Three nuclear groups namely pars posterioris, pars dorsalis, pars ventralis were identified in preoptic area. The inferior lobes are massive and consist of five circumscrible nuclear complexes. Midbrain consists of optic tectum, torus longitudinalis and tegmentum where different nuclear groups were identified.
doi : 10.5214/ans.0972.7531.1118303
Introduction
The teleosts are a highly heterogeneous group of vertebrates with over 31,500 species. Pisces group show a large number of morphological variation. These morphological variations have arisen due to diverse environmental conditions. Although, there is some information available on the cytoarchitectonic pattern of the brain of certain fishes like Sea bass, Clarius batrachus, Notopterus notopterus, Carassius auratus and Cyprinus carpio1-5 but there is hardly any literature available on the snake headed fish Channa marulius. Here in, we describe the cytoarchitectonic pattern of the brain of the giant snake headed murrel, Channa marulius belonging to the family Channidae. This teleost fish can withstand extreme environmental conditions which becomes a unique feature related to habitat. The murrel is much neglected in neuroanatomical terms and their study is necessary step in tracing the evolutionary trends. Hence, an attempt has been made to identify the various nuclear groups from the entire brain region on the basis of standard cytoarchitectonic criteria, to reveal the cytomorpholgy of constituent cell and to study the interrelationship with a view to analyze the overall organization in the light of available literature. The cytoarchitectonic pattern of the forebrain, midbrain and hindbrain of Channa marulius is explored using standard techniques at light microscopic level.
Methods
Channa marulius commonly inhabits fresh water ponds, irrigation tank and rivers. They can thrive in swamps, weedy marshes and can withstand slightly brackish water. It is a carnivorous bottom feeder, feed on variety of animals. The breeding season is from April to June. Fishes were collected from natural habitat all around the Amravati city. They were brought to the laboratory and were acclimatized in aquarium. Twelve adult murrels, Channa marulius of both sexes weighing between 100 to 150 gm and length 14 to 18 cm were used in the present study. Fishes were anaesthetized with 0.2% in 2-phenoxy ethanol and perfused transcardially with 150-200 ml ice cold phosphate buffered saline (PBS, pH-7.45), fol-lowed by the same volume of ice cold Bouin’s fixative. The brain tissue was dissected and fixed in Bouin’s fixative for 24 hours, dehydrated in graded series of alcohol and embedded in paraffin after clearing in xylene. Serial sections of the brain were cut in transverse and saggital planes at 8-10µm thickness. No ethical clearance was required as this does not fall under the purview of Institute ethical committee guidelines.
Staining
Cresyl violet acetate stain was performed by using 1% Cresyl violet (Sigma#C-5042) aqueous solution followed by required dehydration and the slides were finally mounted in DPX as per established procedures.6,7
The deparaffinised section of brain were passed through descending grades of alcohol, brought to water and then stained in haematoxylin. The sections were passed through ascending grades of alcohol up to 70% alcohol and then stained in Eosin in 70% alcohol, dehydrated and mounted in DPX after clearing in xylene.6
Results
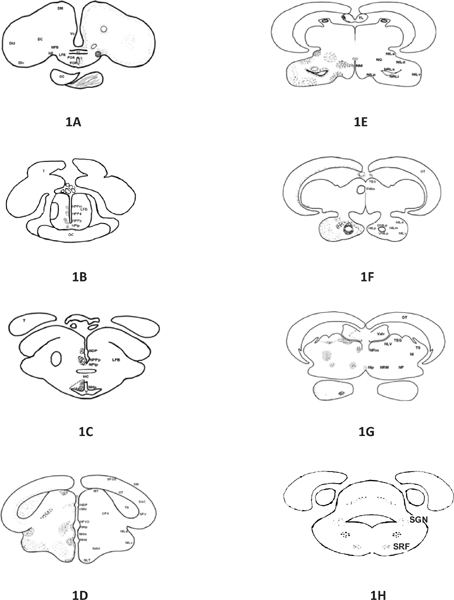
The brain of Channa marulius is divided into forebrain, midbrain and hindbrain. Fig 1 (A-H) show transverse sections through different brain regions (forebrain, midbrain, and hindbrain) of fish Channa marulius. The various areas and neuronal groups in the brain of Channa marulius were identified on the basis of cytoarchitectonic features described in earliear studies of other teleosts.2,3
Forebrain
Channa marulius, forebrain is divided in to telencephalon and diencephalon
Telencephalon
Telencephalon is divisible into two main regions i.e., area dorsalis telencephali and area ventralis telecencephali. Organization of forebrain is depicted in Fig 1A and 2 A-H. The area dorsalis telencephali is subdivided into pars medialis (Dm), pars centralis (DC), pars lateralis dorsalis (Dld) and pars lateralis ventralis (Dlv), depicted in Fig 2A-E). The neurons are rounded oval and triangular in shape, small in size, located near the medial region of the dorsal telencephalon (Dm) and show moderate Cresyl violet staining. The neurons in DC area are sparsely distributed and show intense Cresyl violet staining. The cells of Dld region are larger in size as compared to Dc and Dm. They appear round in shape and exhibit intense Cresyl violet reactivity. While neurons in the area Dlv are compactly arranged, round shape, small in size and are Cresyl violet positive. The area ventralis telencephali is broadly divided into pars ventralis (Vv), pars dorsalis (Vd), pars lateralis (Vl) and pars supracommissuralis (Vs). The cells of Vd are present in groups, round in shape and brightly stained with Cresyl violet staining (Fig 2G). The neurons of the Vv are observed in clusters with small size and are round in shape with intense Cresyl violet staining. The cells of the Vl are rounded in shape, small in size and also show intense staining with Cresyl violet. In Vs region elongated fibers are brightly stained in Cresyl violet preparations (Fig 2H). The lateral forebrain bundles are present in ventro lateral position of the telencephalon as well as medial forebrain bundle (MFB) is present in ventro medial position. The neurons adjacent to the forebrain bundle bigger sized unipoler neurons are also seen forming a cluster which is nucleus entopeduncularis (NE) (Fig 2F) and are Cresyl violet positive. Telencephalic lobes are connected with each other by anterior commissure (AC) (Fig 2H) which is located to the preoptic recess.
Fig. 1: A-H: Schematic drawings of a rostrocaudal series of transverse sections (A-H) of the brain of Channa marulius showing, area dorsalis telenXcephali/pars centralis; area dorsalis telencephali/pars lateralis dorsalis ; Area dorsalis telencephali/pars lateralis ventralis ; Dm, area dorsalis telencephali/pars medialis; AC, anterior commissure ; LFB ,lateral forebrain bundle;MFB,medial forebrain bundle ; NE,nucleus entopeduncularis; OC, optic chisma; Vd, area ventralis telencephali/pars dorsalis; Vl, area ventralis telencephali/pars lateralis; Vs, area ventralis telencephali/pars supracommissuralis; Vv, area ventralis telencephali/pars ventralis; PVO, paraventricular organ; POA, preoptic area; POR, preoptic recess; T, tuberal area; NHd, nucleus hypothalamicus dorsalis; NPPp, nucleus preopticus periventricularis posterioris; NHp, nucleus hypothalamicus posterior; HC, horizontal commissure; V, ventricle; NDP, nucleus dorsalis posterioris of thalamus; NAH, nucleus arcuatus hypothalamicus; NHm, nucleus hypothalamicus medialis; NILp, Nucleus lobi inferioris posterior; NLId, nucleus lobi inferioris dorsalis; NLIm, nucleus lobi inferioris medialis; NLIv, nucleus lobi inferioris ventralis; NLT, nucleus lateralis tuberis; SAC, stratum alum central; SFGS, stratum griseum et album superficial; SM, stratum marginale; SPV, stratum periventriculare; TS, toruus semicirucularis; OC, optic chisma; NRLs, nucleus recessus lateralis superior; NRM, nucleus reticularis messacephali; NLTa, nucleus lateralis tuberis/pars anterior; NRLi, nucleus recessus lateralis inferior; TL, torus longitudinalis; Valv, valvula cerebella; VNLIp, vevtral nucleus lobi inferioris posterior; SGN, secondary gustatory nucleus; SRF, superior reticular formation.
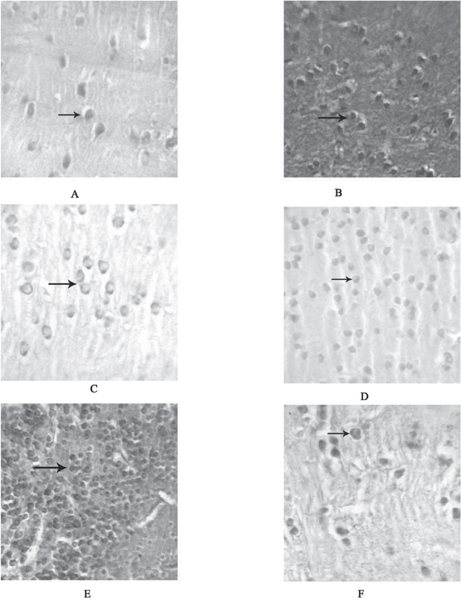
Fig. 2: (A-Q) – Photomicrograph of the transverse section through the brain regions of Channa marulius showing different neuronal complexes. (A) Transverse section of the brain through telencephalon showing intense staining of cresyl violet in neurons of pars centralis (DC) (arrow) 100X. (B) Transverse section of the brain through telencephalon showing intense staining of Haematoxylin - Eosin is seen in neurons of pars centralis (DC) (arrow) 40X. (C) Transverse sections of the brain through telencephalon showing pars lateralis ventralis (Dlv) (arrow) with positive staining of cresyl violet 100X. (D) Transverse section of the brain through telencephalon showing moderate staining of cresyl violet in cells of nucleus of pars medialis (DM) (arrow) 40X. (E) Transverse section of the brain through telencephalon showing pars lateralis dorsalis (Dld) (arrow) showing intense staining of cresyl violet 40X. (F) Transverse section of the brain through telencephalon showing Nucleus entopeduncularis (NE) (arrow) with positive staining of cresyl violet 100X.
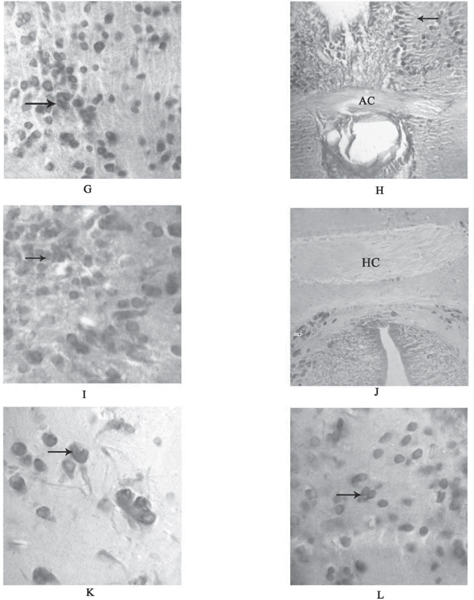
Fig. 2: (G) Transverse section of the brain through telencephalon showing pars dorsalis (Vd) (arrow) brightly stained with cresyl violet 100X. (H) Transverse section of the brain through telencephalon showing pars supracommissuralis (VS) (arrow) elongated fibers was brightly stained in cresyl violet preparations at the level of anterior commissure (AC) 40X. (I) Transverse section of the brain through diencephalon showing nucleus of Torus longitudinalis (TL) (arrow) were brightly stained in cresyl violet 40X. (J) Transverse section of the brain through diencephalon showing Horizontal commissure (HC) (arrow) was brightly stained in cresyl violet preparations 40X. (K) Transverse section of the brain through inferior lobe area showing Nucleus lobi inferioris posterior (NLIp) (arrow) positively stained with cresyl violet 100X. (L) Transverse section of the brain through inferior lobe area showing Nucleus lobi inferioris dorsalis (NLId) (arrow) positively stained with cresyl violet 40X.
Diencephalon
Diencephalon is divided into thalamus, epithalamus and hypothalamus. Hypothalamus is divided in to preoptic area, postoptic area, and tuberal area. The nuclear complexity of the hypothalamic subdivisions is mostly along the periventricular region. Caudal to the optic chiasma, the tuberal area is present, which is demarcated by the commencement of horizontal commissure (HC). A distinguishable laterally directed diverticulum is the lateral recess formed by the lateral expansion of the third vertical. The dorsocaudal edge of the hypothalamus is separated from the posterior tuberculi by the paraventricular organ. The oval shaped inferior lobes are laterally expanded on the either side of the tuberal area. The lateral recess enters and curves within the inferior lobes.
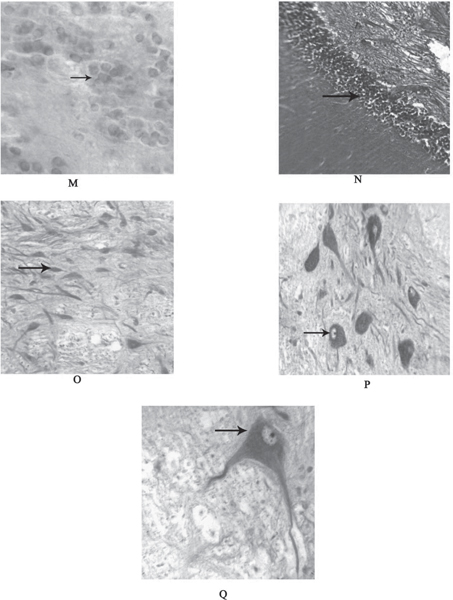
Fig. 2: (M) Transverse section of the brain through tuberal area showing Nucleus arcuatus hypothalamicus (NAH) (arrow) with cresyl violet 100X. (N) Transverse section of the brain through rhombencephalon showing Nucleus Superior reticular formation (SRF) (arrow) moderately stained with Haematoxylin-Eosin 40X. (O)-(P) Transverse sections of the brain through rhombencephalon showing unipolar Secondary gustatory nucleus (SGN) (arrow) intense stained with cresyl violet 100X respectively. (Q) Transverse sections of the brain through rhombencephalon showing bipolar Secondary gustatory nucleus (SGN) (arrow) intense stained with cresyl violet 100X.
Preoptic area and Postoptic region
Fig 1B is a line diagram which shows the telencephalic lobes which are reduced and seen on the dorsal side of preoptic processes. Optic chisma (OC) is observed on the ventral side of the diencephalons. Neuronal complexes are observed in pairs on both sides of the preoptic recess (PR). The nucleus preopticus periventricularis ventrilis (NPPv) represent the anterior most neuronal complex of the hypothalamus located on either side of the preoptic recess. Nucleus preoptic periventricularis dorsalis (NPPd) commences at mid level of the anterior commissure, posteriorly the nucleus is placed lateral to the preoptic recess and occupies a position dorsal to the nucleus poreppticus periventricularis ventralis (NPPv). Nucleus preopticus periventricularis medialis (NPPm) seen on the dorsal side of the nucleus preopticus periventricularis dorsalis. Nucleus preopticus periventricularis ventralis (NPPv) cells are observed, which are small in size and round or oval in shape. The cells of nucleus preopticus periventricularis medialis observed are morphologically large in size and moderately stained. The cells observed in this area occur in cluster of 10 to 12 cells and have stained intensely with cresyl violet. Fig 1C depicts preoptic as well as postoptic region of the diencephalon. It also shows the emergence of optic tectum (OT) from the dorsal side. Horizontal commissure (HC), hypothalamus is observed in the ventral side of the tuberal area and at the caudal side of the Horizontal commissure (HC) (Fig 2J). In the anterial tuberal area, third ventricle is observed and around the third ventricle a group of complex neurons was observed, which are Nucleus Hypothalmicus (NH). In the postoptic area, a prominent area, scattered nuclei have been identified viz. nucleus postopticus lateralis (NPlp). The nucleus postopticus lateralis commences (NPlp) ventrolaterally to nucleus preopticus periventracularis posterior (NPPp) depicted in Fig 1C. The neurons are isolated, round or oval in shape and moderately stained with cresyl violet.
Tuberal Area
In the anterior tuberal area around the third ventricle, three prominent nuclei can be distinguished viz. nucleus hypothalamicus posterialis (NHp), nucleus hypothalamicus dorsalis (NHd), and nucleus hypothalamicus medialis (NHm). The nucleus hypothalamicus posterialis (NHp) consists of 2-3 cells thick layer of neurons located around, slit – like, on either side of the third ventricle, (Fig 5) that commences posterior to the horizontal commissure. The cells are brightly stained, round or oval in shape and smaller in size, with moderate amount of cytoplasm and show small round nuclei. Nucleus hypothalamicus medialis (NHm) consist of 1 to 2 cell thick layer, brightly stained, situated in between the nucleus hypothalamicus posterior (NHp) and nucleus hypothalamicus dorsalis (NHd). The nucleus hypothalamicus dorsalis is situated dorsal to the nucleus hypothalamicus medialis. The neurons are small and intensely stained with cresyl violet and are round or oval in shape (Fig 1 C-D). Nucleus anterior tuberis (NAT) is large group of neurons located in the mid-tuberal area, lateral to the nucleus hypothalamicus posterior. In the ventral tuberal area, the nucleus lateralis tuberis (NLT) is prominent nucleus located at ventromedial aspect (Fig 1D). It commences at the level of the HC and extends caudally as far as pituitary gland.
The nucleus lateralis tuberis is divisible into two parts; nucleus lateralis, tuberis pars anterior (NLTa) and nucleus lateralis tuberalis pars posterior (NLT). The cells of nucleus lateralis tuberalis pars anterior (NLTa) are situated close to ependymal border of the third ventricle. The nucleus lateralis tuberalis posterior is situated immediately behind the nucleus lateralis tuberalis (NLTa) and possesses mostly unipolar cells which are smaller and show round in shape and intensly stained nuclei. Nucleus arcuatus hypothalamicus (NAH) (Fig 2M) is inverted in the caudal tuberal area. Caudal to the tuberal area, lateral side of the third ventricle shows lateral recessus. Around the lateral recess, compactly arranged cells of nucleus recessus lateralis (NRL) are observed (Fig 1E). This nucleus is subdivided on the basis of its position into nucleus recessus lateralis superior (NRLs) and nucleus reecssus lateralis inferior (NRLi). Nucleus recessus lateralis superior consists of compactly arranged spindle shaped cells and nuclei that are clearly seen and show intense cresyl violet stain. Nucleus recessus lateralis interior (NRLi) cells are spindle shaped and nuclei can be seen and show intense cresyl violet stain.
In the transition zone between ventral thalamus and interior lobes, isolated nuclear group have been seen, i.e. nucleus glomerulosus (NG) (Fig 1E). The neurons are moderately sized and oval or round in shape and moderately cresyl violet stained. The caudomedial area includes the nuclei of nucleus mammilliaris (NM) (Fig 1E). The nucleus mammilliaris are very small and area is like pouch and weakly cresyl violet stained. The nucleus of the paraventricular organ (NPVO) is located slightly lateral to the paraventricular organ (PVO) and extends dorsalwards (Fig 1D). The neurons are small and are weakly cresyl violet stained.
Inferior Lobe
The inferior lobes are massive and consist of seven clearly circumscribable nuclearcomplexes. Nucleus lobi inferioris dorsalis (NLId) (Fig 2L) begins at rostral end of the inferior lobes and continuous dorsally (Fig 1E- F) in transverse section, the nuclear group assumes oblong outline although several neurons are small; they are intermixed with smaller cells. Irrespective of their size, cells are round or polymorphic. Nucleus lobi inferioris anterior (NLIa) is situated in the dorsomedial part of the lobes. It commences at the rostral end and is confined to the anterior one third of the lobes. Nucleus lobi inferioris ventralis (NLIv) is the largest group which occupies the bulk of the lobes. Rostrally, this nucleus is located ventrolaterally region and gradually takes the ventral position. Nucleus lobe inferiors medialis (NLIm) consists of few moderate sized cells dispersed over a considerable area in the core of caudal half of the lobes. Nucleus lobi inferioris porterior (NILp) is confined to the caudal part of the lobes.
Nucleus lobi inferioris posterior (NLIp) (Fig 1F, 2K) is divided into two, one is dorsal nucleus lobi inferioris posterior (DNLIp) and second is ventral nucleus lobi inferioris posterior (VNLIp). The neuronal cells of nucleus lobi inferioris anterior (NILa) are small in size, round or oval in shape and are showed in group of 15-20 cells and also showed moderate cresyl violet stain. The neuronal cells of nucleus lobi inferioris ventralis (NLIv) are moderate in size and round or oval in shape and showed moderate cresyl violet stain. It is the largest neuronal cell group in the inferior lobes, these all observation depicated in Fig 1E.
Midbrain (Mesencephalon)
Optic tectum
Isolated, unipolar, oval or rounded neurons are seen in the stratum gresium et album superficial (SFGS). Very few neurons are observed in the stratum marginali (SM). Stratum marginali is identified as the outermost layer. Located on the continuous ribbon like neuronal cells of 1 to 2 layers are seen in stratum alum centrale (SAC). These neurons are moderate in size and oval or rounded in shape and moderately stained. The latter send their processes right across the tectum. Rounded or oval and intensely cresyl violet stained neurons are seen in stratum periventriculare (SPV) (Fig 1G).
Torus longitudinalis
In the torus longitudinalis (TL), the anterolateral area shows the smaller in size and oval or rounded in shape and compactly arranged neuronal cells. Later on the scattered throughout torus longitudinalis. The neurons are moderately stained by cresyl violet (Fig 1E- F).
Tegmentum
Most of the nuclear groups in mesencephalic tegmentum of Channa marulius were moderately stained with cresyl violet. Ventrally to the optic tectum i.e. torus semicirularis (TS) compactly arranged neurons were seen in torus semicirularis (TS) (Fig 1G). Neurons are smaller in size and rounded or oval in shape and moderately stained with the cresyl violet. In the central pretectal area (CPA), conspicuous groups of 8 to 12 neuron are seen. Neurons are large in size, spindle in shape. Moderately stained cytoplasm and intensely cresyl violet stained nuclei with are seen and also axons. Neurons of (CPA) central pretectal area are unipolar. The dorsal midbrain tegmentum (MT) neuron is seen. The neurons are smaller in size and rounded or oval in shape and moderately cresyl violet stained. Lateral side of the valvula cerebelli shows the nucleus lateralis valvulae (NLV) (Fig 1G), Neuron are smaller in sized, oval or rounded shape, they are observed in cluster and moderate cresyl violet stained. Ventrally to the torus semicirularis nucleus isthmi are observed, showing clustered and moderate size neurons which are spherical in shape. They show moderate cresyl violet staining. Ventral to the valvula cerebelli, nucleus fasciculus longitudinalis medialis (NFLm) (Fig 1G-F) is observed. The neurons are few in number, large in size, spherical in shape, with intense cresyl violet staining. Ventrolateral to the fasciculus longitudinalis medialis, nucleus reticularis mesencephali (NRM) are observed. The neurons are small in size, oval or rounded in shape, scattered and moderate cresyl violet stained.
Ventrally to the nucleus lateralis valvulae (NLV), the nucleus profoundus mesencephali (NP) was distinguished (Fig. 1G). Neurons are few, scattered, smaller in size, round or oval in shape and showed weakly cresyl violet stained. In the caudal part of the tegmentum, nucleus interpenduncularis are observed at the median portion. The neurons are small in size and oval or rounded in shape and showed moderate cresyl violet staining.
Hind brain (Rhombecphalon)
The secondary gustatory nucleus (SGN) is located in the lateral part of the fourth ventricle. The neurons in this region are spindle shaped, large in size, with conspicuous nuclei. Situated on the ventral side of fourth ventricle, 3 – 4 cells of nuclear groups is observed which shows large spherical neurons. Dorsally, the fourth ventricle, semicircular by neuronal cells, with prominent nuclei and axon. All the nuclear groups are moderately stained with cresyl violet.
Discussion
In the brain of Channa marulius, different groups of cells with diverse morphology were located in forebrain, midbrain, and hindbrain regions. The cells occurred in distinct clusters or in isolated form. In Channa marulius the telencephalon, divided into dorsalis telencephali and ventralis telencephali, show the presence of Dm, Dc, Dld, Dlv nuclear groups and Vv ,Vd ,Vl, Vs, Vp , NE respectively. These appear to correspond with the different teleost fishes.3, 4, 6, 8-11
Diencephalon is considered to be the most complex region in the brain of the teleost.12 It is the neuronal integration centre of gustatory information, reproduction and vision in the goldfish.4 Using cytoarchitectonic criteria, epithalamus, thalamus, hypothalamus and preoptic area include as the parts of the diencephalon in Dicentrarchus labrax and rayfinned fish.13-14 This type of division of diencephalon is used in the cytoarchitcetural analysis to study of organization of neuronal groups in the brain of different teleosts.4,8-10,14 Similar pattern of distribution of nuclear groups of diencephalon was observed in Channa marulius. The lateral forebrain bundles (LFB) and medial forebrain bundle (MFB) are present in ventro lateral and ventro medial position respectively in Channa marulius. Adjacent to these forebrain bundles, a cluster of unipolar neurons is observed which is the nucleus entopeduncularis (NE). Whereas the same neuronal group i.e, NE in N. Notop-terus is varying in morphology, showing multipolar neurons.3 These neurons show moderate cresyl violet staining.
Preoptic area is the part of rostral most diencephalic subdivision.11 A dense aggregation of nuclei around the third ventricle was observed in Channa marulius. This is comparable with similar pattern of arrangement in Clarius batrachus and Notopterus notopterus.2,3 In Channa marulius, nucleus postopticus laleralis (Nplp) is ventrolatral to nucleus preopticus periventricularis posterior (NPPp). The nucleus postopticus laleralis (Nplp) is comparable with Nplp of Clarius batrachus.2 In Channa marulius, periventricular zone of the anterior tuberal area consists of nucleus hyphthalamicus dorsalis (NHd), medialis (NHm) and posterialis (NHp). The NHd and NHv seem to be comparable with hypothalamicus dorsalis and hypothalamicus ventralis respectively in Clarius batrachus.2 In goldfish, bichir and longnose gar and in cichlid fish4,15 the dorsal hypothalamic nucleus is described in the similar position.
Another nuclear group, nucleus lateralis tuberis (NLT) has been extensively studied in several teleosts.4,16 In Channa marulius, on the basis of the size of the neurons and their position, the NLT could be divided into two division viz., the NLT-pars anterior and NLT-pars posterior. Similar position of NLT has been reported in Clarius batrachus.2,17 However in Salmon sp.18 NLT has been divided into pars rostralis, medialis, lateralis and ventrolateralis. The NLT neurons were reported to be absent in some species.2
In the caudal part of the tuberal area, on the lateral side of the third ventricle, hypothalamus gives extensions called as the lateral recesses: lateralis superior and inferior in Channa marulius. In salmonids, catecholamine producing cells of nucleus recesses latealis (NRL) and NRL in Clarius batrachus seem to be the similar ones.2 The nucleus paraventricular organ (NPVO) is located slightly lateral to the paraventricular organ (PVO), shown in Clarius batrachus and comparable with the present study.2 The nucleus mamillaris (NM) of Channa marulius seems comparable with the Clarius batrachus.2 In Channa marulius, five prominent nuclei were identified in the inferior lobe of hypothalamus viz., nucleus lobi inferioris dorsalis (NILd), nucleus lobi inferioris anterior (NILa), nucleus lobi inferioris ventral is (NILv), nucleus lobi interioris medialis (NILm) and nucleus lobi inferioris posterior (NILp). Similar findings are noted in Clarius batrachus and Notopterus notopterus.2,3 But earlier workers have shown only one nucleus i.e., nucleus diffuse lobi inferioris.4,19
The midbrain Channa marulius showed extensive distribution of neuronal groups of optic tectum of stratum gresium et album superficiale (SGAS) and stratum periventriculare (SPV). These two nuclear groups were observed to be immunoreactive for CARTp in Clarius batrachus.20 Neuronal cell layers of stratum periventricular (SPV) and stratum alum centrale (SAC) are observed in Channa marulius. These cell layers have been reported in the minar carp B. bendelisis.21 Large numbers of nuclear cells were seen in torus semicirularis (TS) of Channa marulius.The central pretectal area showed large spherical shape neuronal group which are similar to as in Clarius batrachus where they were showed to be CARTp positive.20
In the ventrolateral to valvula cerebella and medial to the torus semicircularis, nucleus lateralis valvulae (NLV) situated in the Channa marulius are similar to that of nucleus lateralis valvulae in the carp, Cyprinus carpio.5 Tegmentum region, in Channa marulius is ventral to the torus semicircularis nucleus isthmi and to the vulvula cerebella, nucleus fasciculus longitudinalis medialis (NFLm). These are comparable to the nucleus isthmi and nucleus fasciculus longitudinalis of the Medaka Oryzias lafipes.22,23 Most of the nucleas in mesencephalic tegmentum of B. bendelisis21 showed moderate presence of SBB and AH positive tests especially in torus semicirularis (TS), nucleus isthmi (NI), nucleus profundas mesencephali (NPM), nucleus occulomotorus (ND) and nucleus trochlearis (NT) where as the concentration of these substances was weak in nucleus interpeduncularis (NIP). All these nuclear areas were observed to be with moderate staining, in HgBPB treatment.24 Similar nuclear group are seen in the Channa marulius except nucleus occulomotorus (ND) and nucleus trochlearis (NT). Four nuclear groups were observed in the hind brain region of Channa marulius. SGN and SRN nuclear groups are comparable with the SGN and SRN of rainbow trout, Oncorhynchus mykiss.25
The article complies with International Committee of Medical Journal Editor’s uniform requirements for the manuscripts.
Competing interests – None
Received Date : 11 March 2011; Revised Date: 6 May 2011
Accepted Date : 3 June 2011
References
1. Cerda – Reverter MJ, Zanuy S and Muñoz-Cueto JA. Cytoarchitectonic study of the brain of a perciform species, the sea bass (Dicentrarchus labrax). I. The telencephalon. J Morphol 2001; 247 (3): 217–228.
2. Ram Krishna NS and Subhedar N. Cytoarchitectonic pattern of the hypothalamus in the catfish, Clarias batrachus (Linn.). J Hirnforsch 1991; 32(3): 289–308.
3. Baile VV, Raut IN and Bhute YV. Organization of olfactory system, forebrain and pituitary gland of a teleost. Notopterus notopterus. Annals of Neurosci 2008; 15(2): 43–50.
4. Peter RE and Gill VE. A stereotaxic atlas and technique for forebrain nuclei of the goldfish, Carassius auratus. J Comp Neurol 1975; 159(1): 69–101.
5. Ito H and Yoshimoto M. Cytoarchitecture and fiber connections of the nucleus lateralis valvulae in the carp (Cyprinus carpio). J Comp Neurol 1990; 298: 385–399.
6. Rodriguez-Gomez FJ, Sarasquete C and Munoz-Cueto JA. A morphological study of the brain of Solea senegalensis. I. The telencephalon. Histol Histopathol 2000; 15(2): 355–364.
7. Rupp B and Northcutt RG. The diencephalon and pretectum of the white sturgeon (Acipenser transmontanus): a cytoarchitectonic study. Brain Behav Evol 1998; 51(5): 239–262.
8. Bhute YV, Masram SC, Raut IN, et al. Cytoarchitectonic pattern of the olfactory system, forebrain and pituitary gland of the Indian major carp, Labeo rohita. Annals Neurosci 2007; 14(2): 30–36.
9. Riedel G. The forebrain of the blind cave fish Astyanax hubbsi(Characidae) I. General anatomy of the telencephalon. Brain Behav Evol 1997: 49 (1): 20–38.
10. Rodrigue-Gomez FJ, Rendon-Unceta MC, Sarasquete C et al. Localization of tyrosine hydroxylase-immunoreactivity in the brain of the Senegalese sole, Solea senegalensis. J Chem Neuroanat 2000; 19(1): 17–32.
11. Nieuwenhuys R, Donkelaar HJ and Nicholson C. In: The central nervous system of vertebrates, Vol. 2. Berlin: Springer 1998; 759–93.
12. Mukuda T. and Ando M. Brain atlas of the Japanese eel: Comparison to other fishes. Mem Fac Integrated Arts and Sci, Hiroshima Univ 2003; 29(IV): 1–25.
13. Cerda–Reverter JM, Zanuy S and Muñoz-Cueto JA. Cytoarchitectonic study of the brain of a perciform species, the sea bass (Dicentrarchus labrax). II. The diencephalon. J Morphol 2001; 247 (3): 229–251.
14. Braford MRJ and Northcutt RG. Organization of the diencephalon and pretectum of the rayfinned fishes. Higher brain areas and functions. In: Davis RE and Northcutt RG Eds., Fish neurobiology, Vol. II, Ann Arbor: Univ. of Michigan Press 1983; 117–163.
15. Fernald RD and Shelton LC. The organization of the diencephalon and the pretectum in cichlid fish, Haplochromis burtoni. J Comp Neurol 1985; 238(2): 202–217.
16. Lazar G, Calle M, Roubos EW, et al. Immunohistochemical localization of cocaine- and amphetamine-regulated transcript peptide in the central nervous system of the frog, Rana esculenta. J Comp Neurol 2004; 477(3): 324–339.
17. Rao PD, Betole UK and Subhedar N. Effect of electrical stimulation of the optic nerve on the nucleus preopticus and nucleus lateralis tuberis of the catfish, Clarias batrachus (Linn.). Neuroendo 1980; 30(4): 232–237.
18. Terlou M and Ekengren B. Nucleus praeopticus and nucleus lateralis tuberis of Salmo salar and Salmo gairdneri: Structure and relationship to the hypophysis. Cell Tissue Res 1979; 197(1): 1–21.
19. Billard R and Peter RE. A sterotaxic atlas and technique for nuclei of the diencephalon of rainbow trout (Salmo gairdneri). Reprod Nutr Develop 1982; 22(1): 1–25.
20. Singru PS, Mazumdar M, Sakharkar AJ, et al. Immunocytochemical localization of Cocaine and Amphetamine-regulated transcript peptide in the brain of the catfish, Clarias batrachus (Linn). J Comp Neurol 2007; 502(2): 215–235.
21. Singh N. Histoenzymological localization of glycerol-3-phosphate dehydrogenase and succinate dehydrogenase in the mesencephalon of a minor carp, Barilius bendelisis (Hamilton). Acta Ichtayol Piscat 1988; 18 (2): 103–110.
22. Kapsimali M, Bourrat F and Vernier P. Distribution of the orphan nuclear receptor Nurr1 in Medaka (Oryzias latipes): cues to the definition of homologous cell groups in the vertebrate brain. J Comp Neurol 2001; 431(3): 276–292.
23. Anken R. and Bourrat F. In: Brain atlas of the Medakafish: Oryzias latipes. Paris: INRA 1998; 1–19.
24. Singh N, Singh HR and Tandon KK. Histochemical mapping of lipids and proteins in the mesencephalic nuclei and fibre tracts of hillstream teleost, Barilius bendelisis (Hamilton). Acta Ichtayol Piscat 1989; 19(2): 97–106.
25. Folgueira M, Anadon R and Yanez J. Experimental study of the connections of the gustatory system in the rainbow trout, Oncorhynchus mykiss. J Comp Neurol 2003; 465(4): 604–619.