Annals of Neurosciences, Volume 19, Issue 4 (October), 2012
Effects of estrogen on the serotonergic system and calcitonin gene-related peptide in trigeminal ganglia of rats
KEY WORDS
Ovariectomy
17β-estradiol
Serotonin
Migraine
ABSTRACT
Background: The prevalence of migraine is 3-folds higher in females than in males, and it is intricately related to the levels of estrogen. Estrogen may regulate the expression of metabolic enzymes and receptors of serotonin and also calcitonin gene-related peptide (CGRP), which are implicated in migraine pathogenesis.
Purpose: To study the effects of estrogen on the components of serotonin system and CGRP in trigeminal ganglia of ovariectomized (OVX) rats.
Methods: OVX rats were administered estrogen in silastic tubes and after 48 h, serum estrogen levels were determined. Trigeminal ganglia tissues were used for RT-PCRs of tryptophan hydroxylase (TPH), monoamine oxidase (MAO), serotonin receptors (5-HT1A, 5-HT1B, 5-HT2A), estrogen receptor (ER) and CGRP. Western blots of TPH and MAO were performed.
Results: Estradiol administration to OVX rats increased TPH mRNA levels, while decreased MAO mRNA levels in trigeminal ganglia tissue. Western blot data correlate with the gene expression results. The decreased mRNA levels of serotonin receptors following ovariectomy were restored in estrogen-replenished rats. The induced gene expression of ER in OVX rats was restored following estrogen replenishment.
Conclusion: Estrogen levels affect the levels of serotonin metabolizing enzymes and its receptors besides CGRP levels. Since TPH and MAO levels regulate circulating and physiologically available serotonin content, the regulation of serotonin metabolizing enzymes suggest a plausible mechanism by which estrogen alleviates migraine in women.
doi : 10.5214/ans.0972.7531.190403
Corresponding Author:
S. Puri, PhD
Tel : +91 172 2534135;
E-mail : spuri_1111@yahoo.com
Introduction
The prevalence of migraine in women has been estimated to be as high as 25%, which is about 3-folds the rate of men.1 Several lines of evidence suggest an association between migraine and reproductive function. A gender difference in migraine incidence2 emerges at adolescence when sex hormones come into play. Migraine is associated with the perimenstrual time period (two days before to three days after the onset of menstruation), the postpartum period and the perimenopause state, all of these are physiological states in reproductive life span of a women characterized by declining estrogen concentrations.3,4 The incidence of migraine is increased during pill-free period of combined hormonal contraception5 and cyclical hormone replacement therapy i.e. following estrogen withdrawal. These observations support role of estrogen level depletion as a trigger for migraine. It is suggested that sex steroids modulate susceptibility in migraine patients by altering the expression of various genes that are implicated in a migraine attack.6 Estrogen may exert rapid non-genomic effect or chronic genomic effect. Welch et al.7 and Puri et al.8 have studied the effects of estrogen on trigeminal ganglia in culture and brain of mice respectively and listed the regulation of various such genes with potential relevance to migraine.
Estrogen acts as a neurosteroid influencing the pain pathway associated with migraine by binding to its receptors (estrogen receptors, ERs) present on the trigeminal nerves.9 The trigeminovascular system consists of a network of cranial vessels and their trigeminal innervations that convey the pain information to the central nervous system where migraine pain is perceived.10 The pain response increases by the release of vasodilating neuropeptides especially calcitonin gene-related peptide (CGRP). Decreased levels of a neurotransmitter, serotonin, are also linked to migraine. The implication of serotonin in migraine pathogenesis has been indicated by studies showing the presence of its receptors (5-HT receptors) on the trigeminal nerve and cranial vessels and also by the relief obtained after serotonin administration. During the regulation of serotonergic neurotransmission, some of the serotonin receptors may be involved in triggering migraines (eg. 5-HT1C)11 and others in preventing them (eg. 5-HT1D).12 Many effective anti-migraine drugs used currently are serotonin receptor agonists such as triptans.
Accordingly, triptans act on trigeminovascular system and bring the elevated serum levels of CGRP to normal13 to bring about migraine relief. The varying levels of estrogen during menstrual cycles lead to alterations in the levels of serotonin-metabolizing enzymes modulating serotonin14 and CGRP levels.15
Estrogen administration have been reported to upregulate tryptophan hydroxylase mRNA (TPH, serotonin synthesizing enzyme),9,16 downregulate serotonin reuptake transporter (SERT) mRNA following short-term treatment17 and upregulate its mRNA following long-term treatment,18 decrease gene expressions of monoamine oxidase (MAO, serotonin catabolizing enzyme)19 and 5-HT1A receptor within dorsal raphe. Yang et al20 showed reduced CGRP levels following estrogen treatment in the rat dorsal root ganglia.
However, the effect of estrogen on the serotonergic system and CGRP levels in the trigeminal ganglia of female rats has not been ascertained. Keeping this in mind, the present work has been designed to study the effect of estrogen on CGRP expression and the serotonin metabolizing enzymes and its receptor expression.
Methods
Animal groups
Three month old female wistar rats (n = 21; weighing 150-200 g), procured from the Central Animal House, Panjab University, Chandigarh, India, were housed under a standard light and dark cycle and given free access to food and water. The animal care and experimental protocols were in accordance with Institutional Animal Ethics Committee (IAEC), Panjab University, Chandigarh. Bilateral ovariectomy or a sham surgery was performed by standardized procedure. The animals were anaesthetized by ketamine hydrochloride and xylocaine as per the approved recommendations. The fur was shaved off and a dorsal midline skin incision was made on both sides to remove the ovaries except in the sham group. The ovariectomized (OVX) animals were placed individually in separate cages and observed for a period of fourteen days for any mortality or disease as well for depletion of any endogenous estrogen.
One group of the OVX rats was administered estrogen by subcutaneously implantation of silastic tubes (Dow Corning, Midland, Michigan) containing 0.1% 17β-estradiol in ethanol. All other groups received the vehicle only. Implants were inserted subcutaneously into the back of the animal under anaesthesia and the animals were used for experimentation immediately after 48 h of implantation.
The study was performed on four groups (n = 5-6 each) of rats viz.: (i) cycling (control), (ii) sham-operated, (iii) OVX, and (iv) estrogen-replenished ovariectomized rats.
Serum 17β-estradiol measurement
Blood samples (800 mL to 1 mL) were collected from the eye vein of all groups of rats. Serum was collected from the blood by centrifugation at 2,000xg for 20 min and serum estradiol level was measured by microparticle enzyme immunoassay (MEIA) kit obtained from Abbott Laboratories Diagnostic Division, Abbott park, USA.
RNA isolation from trigeminal ganglia tissue
Trigeminal ganglia from rats of all the four groups were harvested and homogenized in TRIZOL Reagent (Invitrogen, CA, USA). Total RNA was extracted following manufacturer’s specifications, precipitated and dissolved in RNase-free water. Integrity of RNA was tested by viewing denatured ethidium bromide stained samples in 1% agarose/formaldehyde gel. Yield and purity of the isolated RNA was spectrophotometrically determined by calculating the ratio of 260/280.
RT-PCR
For semi-quantitative determination of mRNA levels, separate RT-PCRs for the specific gene product and housekeeping gene Rig/S15 were performed as directed by the manufacturer using superscript III one-step RT-PCR system (Invitrogen). Metabion International AG (Deutschland) synthesized the primers. Primer sequences, predicted product sizes and the number of cycles for amplification are provided in Table 1.
Each reaction was run for 30 min at 55°C for 1 cycle and 2 min at 94°C for 1 cycle for cDNA synthesis, followed by cycles of 1 min at 94°C, 1 min at 55°C and 1 min at 68°C for 25 to 50 cycles, and finally 5 min at 68°C for 1 cycle in a Master Cycler Gradient (Eppendorf). The cycle number for each primer set was optimized empirically by determining the number of cycles to reach the plateau phase and by carrying out hybridizations within the linear range of amplification. The amplification products were visualized by electrophoresis using 2% ethidium bromide-stained agarose gel and analyzed using Scion Image software (Scion Incorporation, USA). A ratio of the gene of interest to the housekeeping gene, Rig/S15 was determined for each sample. Ratios were averaged to obtain mean gene levels and differences were assessed.
Table 1: Primer sequence
| Gene | Primer Sequences (5’ -sense-3’; 5’ -antisense-3’) |
Product Size in bp | No. of PCR Cycles |
|---|---|---|---|
| ER | 5’ -AAG AGA AGG ACC ACA TCC ACC- 3’ 5’-GGA ATG TGC TGA AGT GGA GC- 3’ |
326 | 25 |
| TPH2 | 5’-TAA ATA CTG GGC CAG GAG AGG-3’ 5’-GAA GTG TCT TTG CCG CTT CTC-3’ |
132 | 37 |
| MAO | 5’-TGG GAA GAT TCC AGA GGA TG-3’ 5’-GCT GAC AAG ATG GTG GTC AA-3’ |
150 | 50 |
| 5-HT1A | 5’-AGC TTA GGA ACT TCG TCG GCA-3’ 5’-CAG AGG AAG GTG CTC TTT GG-3’ |
200 | 37 |
| 5-HT1B | 5’-GTT GAC TTG TCA ATG GCA T-3’ 5’-GCT TCA GTT CAC ATT CCA GA-3’ |
200 | 36 |
| 5-HT2A | 5’-AGC CGC TTC AAC TCC AGA A-3’ 5’-TTT TGC TCA TTG CTG ATG GA-3’ |
410 | 35 |
| CGRP | 5’-TCC TGC AAC ACC GCC ACC TG -3’ 5’-GGT GGG CAC AAA GTT GTC CT -3’ |
90 | 33 |
| Rig/S15 | 5’-TTCCGCAAGTTCACCTACC-3’ 5’-CGGGCCGGCCATGCTTTACG-3’ |
361 | 30 |
Western blot analysis
In brief, ganglia tissues were homogenized using lysis buffer consisting of 10 mM Tris HCl buffer (pH 7.4) containing 1% Triton X-100, 0.1% SDS and 150 mM NaCl. Protease inhibitors, 1 mM phenyl methyl sulfonyl fluoride, 2 mM sodium orthovanadate, 5 mM EDTA, 2 mM EGTA and 5 µM pepstatin A were added to the lysis buffer. The tissue lysates were centrifuged at 12,000×g for 5 min at 4°C. The clear lysates were used for western blots. Prior to western blot analysis, protein content was determined by the method of Lowry et al.21 Samples containing 20 mg proteins were separated in 12% SDS-polyacrylamide gels and electrophoretically transferred to polyvinylidene fluoride membranes (Milipore). Membranes were incubated in blocking buffer containing 5% skim milk in TBS with 0.1% Tween-20 and then probed with primary antibody (anti-TPH antibody, #sc-30079 and anti-MAO antibody, #sc-50333, Santa Cruz Biotechnology, Santa Cruz, CA, USA) in blocking buffer overnight at 1:500 dilution. Membranes were rinsed in TBS-Tween-20 and incubation with alkaline phosphatase conjugated anti-rabbit secondary antibody (#A3812, Sigma Aldrich) was done at 1: 15,000 dilution in blocking buffer for 45 min at room temperature. CDP-Star detection reagent (GE Healthcare, Amersham, UK) was used for chemiluminescent detection of alkaline phosphatase according to manufacturer’s instructions. Blots were reprobed with 1:15,000 anti-β-actin antiboby (#sc-130657, Santa Cruz Biotechnology, Santa Cruz, CA, USA) and normalized to verify equivalent protein loading. Differences in protein immunoreactivity between treatments were determined by scanning densitometry in proportion to β-actin immunoreactive bands.
Statistical analysis
Data was represented as mean + standard error (n = 5-6). Statistical analysis was performed by using one-way analysis of variance (ANOVA) followed by post hoc test for ascertaining significant difference. Statistical significance of the results was calculated at P<0.05.
Results
Serum 17β-estradiol levels
The levels of 17β-estradiol depleted significantly (P<0.05) on day 15 i.e. after ovariectomy (14.71 + 2 pg/mL) as compared to cycling animals (35.00 + 4 pg/mL) (Table 2). After 48 h of silastic tube implantation in the ovariectomized rats, serum 17β-estradiol levels were significantly increased (68.43 + 2 pg/ml, P<0.05) in comparison to OVX or cycling animals. In sham-operated animals, estrogen concentration was 30 + 1 pg/ml which was not significantly different from that of control group.
Effect of 17β-estradiol on serotonin metabolizing enzymes
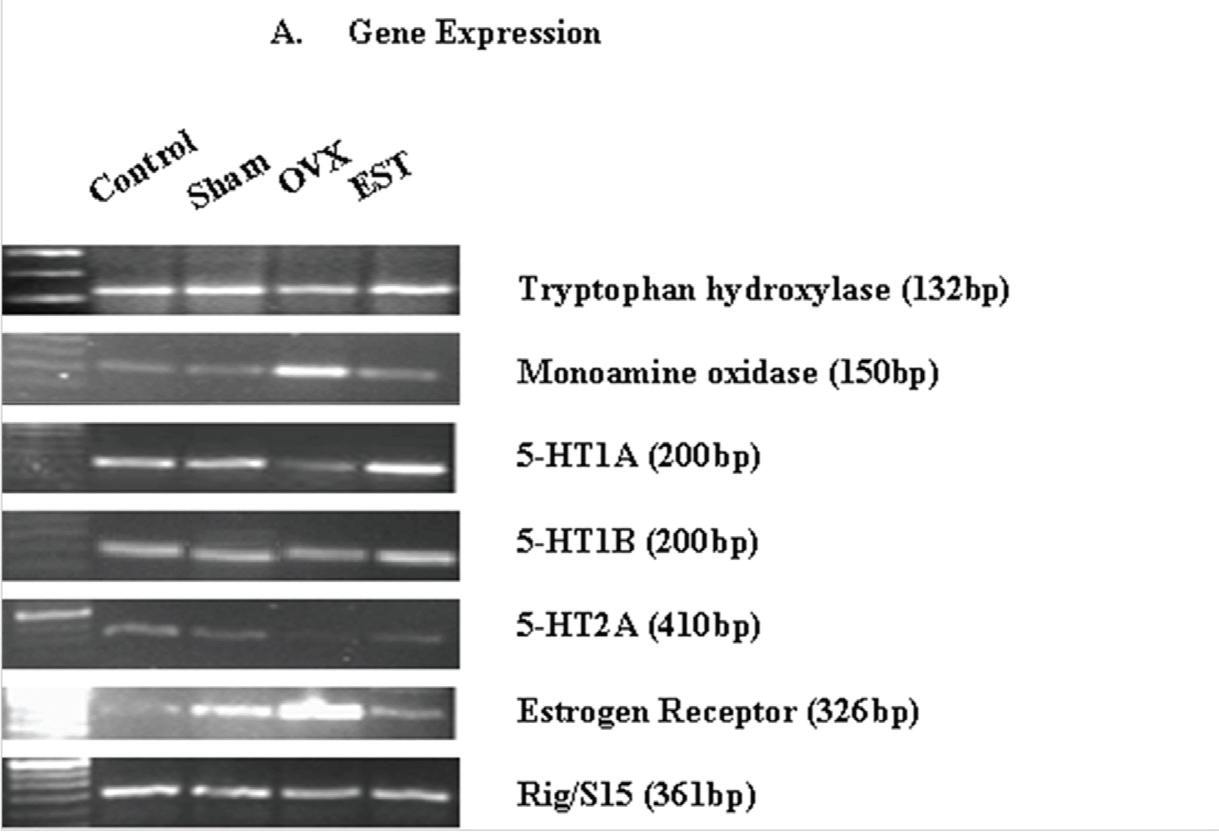
Tryptophan hydroxylase is the rate limiting enzyme in the synthesis of serotonin22 which is degraded mainly by the action of monoamine oxidase enzymes.23 Our results demonstrated that estradiol increases TPH mRNA levels while it decreases MAO mRNA levels in trigeminal ganglia of wistar rats. Fig. 1a demonstrates the gene expression profile of TPH and MAO in trigeminal ganglia of cycling (control), sham-operated, ovariectomized and estrogen-replenished ovariectomized rats. Quantitation of RT-PCR data (mRNA ratios of target genes to housekeeping gene, Rig/S15; Fig. 1b) demonstrates significant decrease of 30% (P<0.05) in TPH gene expression while a significant (P<0.05) 4-fold increase in MAO gene expression following ovariectomy as compared to the cycling rats. Sham-operated animals did not show any significant change as compared to cycling rats. Administration of estradiol to the ovariectomized rats restored gene expression of TPH and lowered down gene expression of MAO to that in control cycling rats. The effects of estradiol at the protein levels of these two metabolizing enzymes correlated with the effects observed at the level of mRNA expressions. The results in fig. 2 demonstrate the western blot analysis of lysate from trigeminal ganglia. Two bands corresponding to ~55kd and ~66 kd were resolved for TPH and MAO respectively (Fig. 2). There was a significant decrease of 20% (P<0.05) in TPH protein expression while a significant increase of 60% (P<0.05) in MAO protein expression following ovariectomy as compared to cycling rats. Sham-operated animals did not show any significant change as compared to cycling rats. Treatment of OVX animals with estradiol increased the amount of TPH protein (40%, P<0.05) and reduced MAO protein levels by 40% (P<0.05) as compared to OVX animals.
Table 2: Serum 17β-estradiol levels in different animal groups
[Values are mean + standard error (n = 5-6).]
| Animal Group | Serum 17β-estradiol levels (in pg/mL ) |
|
|---|---|---|
| i. | Cycling rats (Control) | 35.00 + 4 |
| ii. | Sham-operated | 30.00 + 1 |
| iii. | Ovariectomized rats (OVX) | 14.71 + 2* |
| iv. | Estrogen-replenished ovariectomized rats (OVX+EST) |
68.43 + 2*$ |
*Means differ from control, P<0.05
$Means differ from OVX, P<0.05
Effect of 17β-estradiol on receptors
We examined gene expression of receptors for serotonin and estrogen in the trigeminal ganglia of OVX and estrogen-replenished rats using RT-PCR. Fig. 1 demonstrates that the receptors- 5-HT1A, 5-HT1B, 5-HT2A and ER, were expressed in rat trigeminal ganglia tissue. Ovariectomy i.e. estrogen deprivation produced a decrease of approximately 50% and 30% in the gene expressions of 5-HT1A and 5-HT1B receptors, respectively as compared to the control rats. Administration of estrogen through silastic implants to OVX rats produced an increase of 150% in the mRNA levels of 5-HT1A as compared to control rats and restored completely the lowered mRNA levels of 5-HT1B receptors. Sham treatment did not show any significant change as compared to the cycling rats. Similarly, 80% decrease was observed in the gene expression of 5-HT2A receptor following ovariectomy as compared to control rats. Estrogen administration produced an increase of 20% (P<0.05) in 5-HT2A receptor gene expression as compared to OVX rats. The increased (approximately 3.5-fold (P<0.05); Fig. 1b) gene expression of ER in OVX rats was restored in the trigeminal ganglia of estrogen-replenished ovariectomized rats.
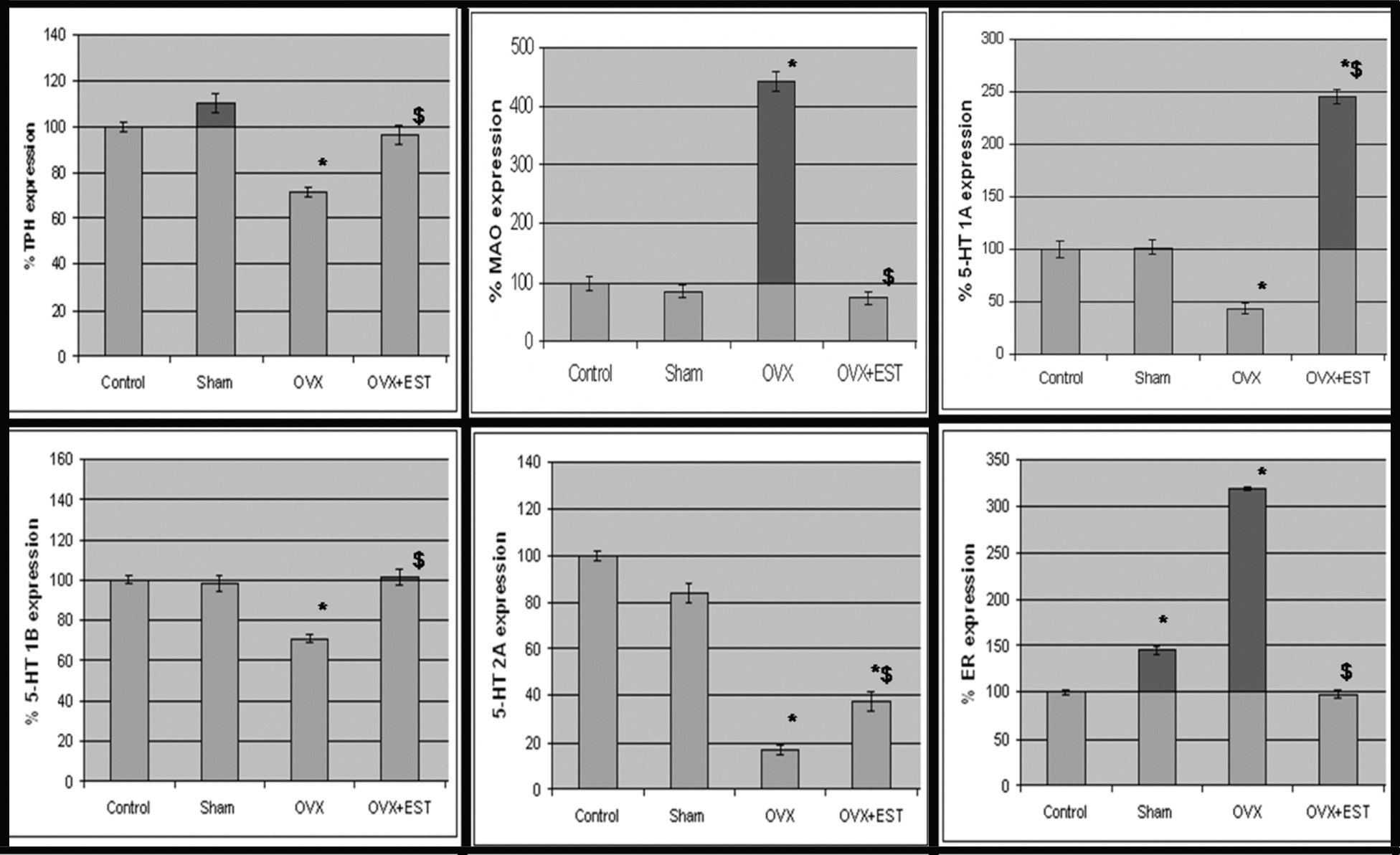
Fig. 1: Expression of genes of the components of serotonin system, estrogen receptor and CGRP in trigeminal ganglia. (a) Ethidium bromide-stained agarose gels of RT-PCRs of tryptophan hydroxylase, monoamine oxidase, 5-HT1A, 5-HT1B, 5-HT2A, estrogen receptor and Rig/S15. Each gel depicts representative data from cycling (control), sham-operated, ovariectomized and estrogen-replenished ovariectomized rats. (b) Quantitation of RT-PCR data demonstrates significant changes in various genes expression. The ordinate represents the ratio of the intensity of the gene of interest to the housekeeping gene (Rig/S15), mean + standard error (n=5). *Means differ from control, P<0.05. $Means differ from OVX, P<0.05.
Effect of 17β-estradiol on CGRP
To determine the effect of estrogen on CGRP in trigeminal ganglia tissue, we assessed the mRNA levels of CGRP in ovariectomized and estrogen-replenished ovariectomized rats. As shown in Fig. 3, ovariectomy significantly increased (P<0.05) the gene expression of CGRP to 600% as compared to control rats. This induced mRNA level was lowered down to 150% following estrogen treatment to the ovariectomized rats.

Fig. 2: Western blot analysis of proteins in trigeminal ganglia of cycling (control), sham-operated, ovariectomized and estrogen-replenished ovariectomized rats. (a) Western blot of TPH, MAO and GAPDH proteins. (b) Quantitaion of proteins using chemiluminescence, mean ± standard error (n=5). *Means differ from control, P<0.05. $Means differ from OVX, P<0.05.
Discussion
Many studies employ multiple subcutaneous injections to ovariectomized animals for estrogen restoration to ensure continuous supply. One major limitation of such studies is that this approach cannot mimic the normal physiological changes in sex hormone levels that occur in intact cycling females.2 Therefore, we have chosen silastic implants that provide a better method for delivering controlled and easily manipulated yet effective dosage of estradiol to ovariectomized rats.24 This ensures stable and sustained estradiol release. It may be noted that OVX rats have a low level of estradiol in the serum. This minimal and basal level is attributed to the production of estradiol by fat and adrenal tissue.25
To assess the effect of ovarian hormone, estrogen, on serotonin neural function, we focused at the level of transcription as well as translation. Our results demonstrated that estradiol increases TPH mRNA expression and decreases the expression of MAO mRNA. Our results are in corroboration with the previous reports of estrogen-induced upregulation of TPH mRNA expression in the dorsal raphe of rodents.14,26 It was observed that the increment in TPH mRNA with estradiol treatment led to a concomitant increase in TPH protein level. It can be tentatively concluded that protein levels of TPH and MAO enzymes reflect their mRNA levels. Overall the increment/enhancement is consistent with ovarian steroids augmenting TPH protein expression9,16 and inhibiting MAO protein expression.19,27 Since TPH and MAO protein levels influence serotonin content, then estradiol might be influencing many other systems further regulated by serotonin. These observations provide a biological basis for the ability of ovarian steroids to alter migraine pain response and other functions regulated by serotonin.
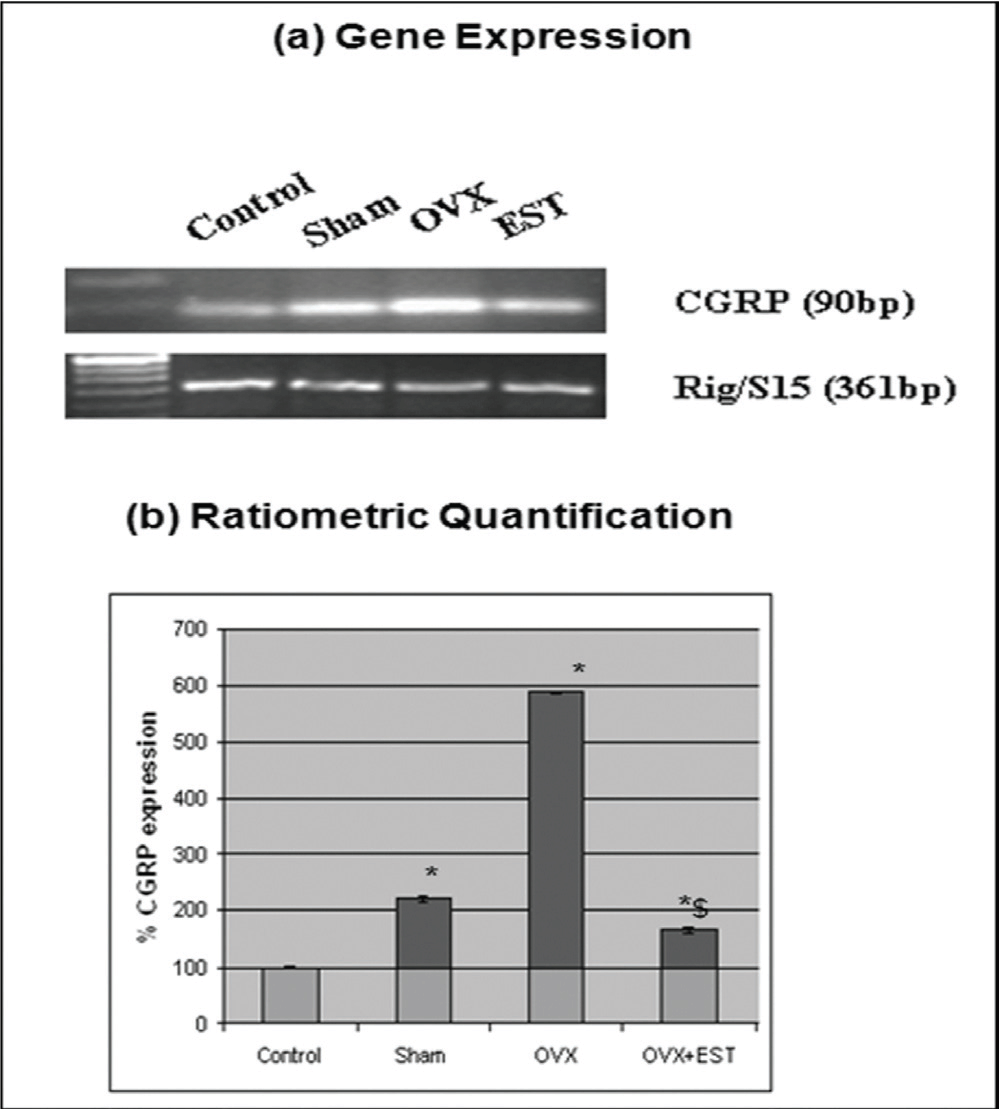
Fig. 3: Expression of CGRP gene in trigeminal ganglia. (a) Ethidium bromide-stained agarose gels of RT-PCRs of CGRP and Rig/S15. Each gel depicts representative data from cycling (control), sham-operated, ovariectomized and estrogen-replenished ovariectomized rats. (b) Quantitation of RT-PCR data demonstrates significant changes in gene expression. The ordinate represents the ratio of the intensity of CGRP gene to the housekeeping gene (Rig/S15), mean + standard error (n=5). *Means differ from control, P<0.05. $Means differ from OVX, P<0.05.
The serotonin receptor system plays a central role in the control of serotonergic neurotransmission and thus features prominently in many behavioral and physiological functions.28 The 5-HT1 and 5-HT2 receptors (especially 5-HT1) have been identified as being responsible for most of the migraine activity.29,30 These receptors are found on the trigeminal nerve endings.31,32 In the present study, we found that the mRNA expressions of 5-HT1A, 5-HT1B and 5-HT2A receptors were decreased following ovariectomy i.e. estrogen deprivation. Administration of estrogen produced an increase in the mRNA levels of 5-HT1A and approximately restored the decreased gene levels of 5-HT1B receptors. Our results are in line with the finding that acute and chronic estrogen treatments increase 5-HT2A receptor mRNA in the dorsal raphe nucleus in ovariectomized rats.33,34 In contrast, it was observed that estrogen treatment resulted in a decrease in 5-HT1A receptor gene expression and receptor function in the limbic region, cortex and dorsal raphe nucleus of rats35,36 and nonhuman primates.37,38 This may be due to dosage or route of administration dependent effect39 of estrogen on the expression of 5-HT1A receptors. The trigeminal ganglia tissue exhibited an increase in the expression of serotonin receptors after estrogen treatment suggesting that the stimulatory effects of serotonin are mediated via 5-HT1A, 5-HT1B and 5-HT2A receptors. Our data suggests that estrogen modulates the density of serotonin receptors and serotonin turnover as is also reported by other studies.40,41 The increased gene expression of ER in OVX rats was lowered down in the trigeminal ganglia of estrogen-replenished rats suggesting negative regulation of ER mRNA expression by estrogen. These results appear consistent with the data indicating estrogen receptor expression is negatively correlated to estrogen levels in trigeminal ganglia sensory neurons42, pituitary cells and cell lines43 and central nervous system tissue.44
CGRP has been implicated in the pathogenesis of migraine through its vasodilatory effect. We observed that ovariectomy increased the gene expression of CGRP which was apparently lowered down following estrogen administration. Our results are in corroboration with studies demonstrating reduced CGRP levels following estrogen treatment in the rat dorsal root ganglia.45 There is evidence of decreased CGRP immunoreactive neurons in the lumbar region of the rat spinal cord following estrogen treatment.45 Elevated CGRP concentrations have been observed in post-menopausal women when estrogen levels decline.46 During attacks of migraine plasma levels of CGRP have been found to be increased in the venous outflow from the head.47,48 CGRP released in the periphery and in the trigeminal brain stem can lead to mast cell degranulation in the dura leading to prolonged activation of meningeal afferents.49 However, Gangula et al.15 and Sarajari and Oblinger50 showed conflicting reports that ovariectomy decreases plasma CGRP concentration in female rats, while subsequent treatment with 17β-estradiol restores it significantly. This may be due to dosage specific or route of administration specific effect of estrogen on CGRP. The increased expression of CGRP during times of lowered estrogen levels hints at a potential mechanism for migraine related to the fall in estrogen concentrations prior to menstruation in affected women.
Conclusion
It could be concluded that estrogen alters the levels of serotonin-metabolizing enzymes and its receptors affecting serotonin levels and also CGRP levels. The experimental model here simulates the physiological conditions of estrogen withdrawal (hormones are depleted), followed by hormonal therapy in human. This condition is frequently concurrent with prevalence of hormonal induced migraine.51 The data provides evidence at transcriptome as well as proteomics level that estrogen regulates serotonin receptors through modulation of serotonin content. This is further being regulated by serotonin metabolizing enzymes. There has been strong evidence that prevalence of migraine is affected by variant expression of serotonin related genes.52,53 These findings suggest that these physiological mechanisms might account for the increased prevalence of migraine headache in women. Further, emerging evidence of the influence of estrogen on serotonergic activity not only provide a mechanistic basis for the association between comorbid situation of migraine and depression rather provides a novelty in drug targets for therapeutic intervention.
Acknowledgements
Funding provided by PURSE (Department of Science and Technology) and Biomedical Sciences to S.P. and University Grants Commission (UGC) to M.A. are thankfully acknowledged. Facilities provided by Centre for Potential with Excellence in Biomedical Sciences, Panjab University, Chandigarh are duly acknowledged.
Abbreviations
CGRP Calcitonin gene-related peptide, RT-PCR Reverse transcription-polymerase chain reaction, 5-HT1A 5-hydroxytryptophan receptor type 1A, 5-HT1B 5-hydroxytryptophan receptor type 1B, 5-HT2A 5-hydroxytryptophan receptor type 2A, ER Estrogen receptor, TPH Tryptophan hydroxylase, MAO Monoamine oxidase, OVX Ovariectomized
The article complies with International Committee of Medical Journal Editor’s uniform requirements for the manuscripts.
Competing interests: None, Source of funding: PURSE.
Received Date : 17 May 2012; Revised Date : 3 July 2012
Accepted Date : 21 September 2012
References
1. Martin VT and Lipton RB. Epidemiology and biology of menstrual migraine. Headache 2008; 48: S124–S130.
2. Cairns BE. The influence of gender and sex steroids on craniofacial nociception. Headache Currents 2007; 47: 319-324.
3. Sances G, Granella F, Nappi RE et al. Course of migraine during pregnancy and postpartum: a prospective study. Cephalalgia 2003; 23: 197–205.
4. Martin VT and Behbehani M Ovarian hormones and migraine headache: understanding mechanisms and pathogenesis- part 2. Headache 2006; 46: 365–386.
5. Aegidius K, Zwart JA, Hagen K, et al. Oral contraceptives and increased headache prevalence: the head-hunt study. Neurol 2006; 66: 349–353.
6. Moskowitz MA. Defining a pathway from bench to bedside: the trigeminovascular system and sensitization. Headache 2008; 48: 688–690.
7. Welch KM, Choudhuri R, Cui L, et al. Estrogen regulation of gene expression in cortical spreading depression: relevance to menstrual migraine. Headache 2002; 42: 440.
8. Puri V, Puri S, Svojanovsky SR, et al. Effects of oestrogen on trigeminal ganglia in culture: implications for hormonal effects on migraine. Cephalalgia 2005; 36: 33–42.
9. Berman NEJ, Puri V, Chandrala S, et al. Serotonin in trigeminal ganglia of female rodents: relevance to menstrual migraine. Headache 2006; 46: 1230–1245.
10. Bolay H, Reuter U, Dunn AK, et al. Intrinsic brain activity triggers trigeminal meningeal afferents in a migraine model. Nat Med 2002; 8: 136–142.
11. Brewerton TD, Murphy DL, Meuller EA, et al. Induction of migraine-like headaches by the serotonin agonist, m-chlorophenylpiperazine. Clin Pharmacol Ther 1988; 43: 605–609.
12. Ferrari MD, Melamed E, Gawel MJ, et al. Treatment of migraine attacks with sumatriptan. The subcutaneous sumatriptan international study group. N Engl J Med 1991; 325: 316–321.
13. Ferrari MD, Roon KI, Lipton RB, et al. Oral triptans (5-HT(1B/1D) agonists) in acute migraine treatment: A meta-analysis of 53 trials. Lancet 2001; 358: 1668–1675.
14. Hiroi R, McDevitt RA, Neumaier JF. Estrogen selectively increases tryptophan hydroxylase-2 mRNA expression in distinct subregions of rat midbrain raphe nucleus: association between gene expression and anxiety behavior in the open field. Biol Psychiatry 2006; 60(3): 288–295.
15. Gangula PR, Lanlua P, Wimalawansa S, et al. Regulation of calcitonin gene-related peptide expression in dorsal root ganglia of rats by female sex steroid hormones. Biol Reprod 2000; 62: 1033–1039.
16. Pecins-Thompson M, Brown NA, Kohama SG, et al. Ovarian steroid regulation of tryptophan hydroxylase mRNA expression in rhesus macaques. J Neurosci 1996; 16: 7021–7029.
17. Pecins-Thompson M, Brown NA, Bethea CL. Regulation of serotonin re-uptake transporter mRNA expression by ovarian steroids in rhesus macaques. Brain Res Mol Brain Res 1998; 53: 120–129.
18. Smith LJ, Henderson JA, Abell CW, et al. Effects of ovarian steroids and raloxifene on proteins that synthesize, transport and degrade serotonin in the raphe region of macaques. Neuropsychopharmacol 2004; 29: 2035–2045.
19. Bethea CL, Lu NZ, Gundlah C, et al. Diverse actions of ovarian steroids in the serotonin neural system. Front Endocrinol 2002; 23: 41–100.
20. Yang Y, Ozawa H, Kawata M, et al. Immunocytochemical analysis of sex differences in calcitonin gene-related peptide in the rat dorsal root ganglion, with special reference to estrogen and its receptor. Brain Res 1998; 791: 35–42.
21. Lowry OH, Rosebrough NJ, Farr AL, et al. Protein measurement with the folin phenol reagent. J Biol Chem 1951; 193: 265–275.
22. Zhang X, Beaulieu JM, Gainetdinov RR, et al. Functional polymorphisms of the brain serotonin synthesizing enzyme tryptophan hydroxylase-2. Cell Mol Life Sci 2006; 63(1): 6–11.
23. Bianchi P, Kunduzova O, Masini E, et al. Oxidative stress by monoamine oxidase mediates receptor-independent cardiomyocyte apoptosis by serotonin and postischemic myocardial injury. Circulation 2005; 112(21): 3297–3305.
24. Cohen PE and Mlligan SR. Silastic implants for delivery of oestradiol to mice. J Reprod Fertility 1993; 99: 219–223.
25. Nelson LR and Bulun SE. Estrogen production and action. J Am Acad Dermatol 2001; 45(3): S116–S124.
26. Charoenphandhu J, Teerapornpuntakit J, Nuntapornsak A, et al. Anxiety-like behaviors and expression of SERT and TPH in the dorsal raphe of estrogen- and fluoxetine-treated ovariectomized rats. Pharmacol Biochem Behavior 2011; 98: 503–510.
27. Ma ZQ, Violani E, Villa F, et al. Estrogenic control of monoamine oxidase A activity in human neuroblastoma cells expressing physiological concentrations of estrogen receptor. Eur J Pharmacol 1995; 284: 71–76.
28. Meneses A. Physiological, pathophysiological and therapeutic roles of 5-HT systems in learning and memory. Terapeutica Experimental Rev Neurosci 1998; 9(4): 275–289.
29. Schmuck K, Ullmer C, Kalkman HO, et al. Activation of meningeal 5-HT2B receptors: an early step in the generation of migraine headache? Eur J Neurosci 1996; 8(5): 959–967.
30. Hamel E. The biology of serotonin receptors: focus on migraine pathophysiology and treatment. Can J Neurol Sci 1999; 26(3): S2-S6.
31. Moskowitz MA and Buzi MG. Neuroeffector functions of sensory fibres: implications for headache mechanisms and drug actions. J Neurol 1991; 238: S18–S22.
32. Bruinvels AT, Landwerhmeyer B, Moskowitz MA, et al. Evidence for the presence of 5HT1B receptor messenger RNA in neurons of the rat trigeminal ganglia. Eur J Pharmacol 1992; 227: 357–359.
33. Sumner BEH and Fink G. Effects of acute estradiol on 5-hydroxytryptamine and dopamine receptor subtype mRNA expression in female rat brain. Mol Cellular Neurosci 1993; 4: 83–92.
34. Sumner BEH and Fink G. Estrogen increases the density of 5-hydroxytryptamine(2A) receptors in cerebral cortex and nucleus accumbens in the female rat. J Steroid Biochem Mol Biol 1995; 54(1–2): 15–20.
35. Osterlund MK and Hurd YL. Acute 17 beta-estradiol treatment down-regulates serotonin 5HT1A receptor mRNA expression in the limbic system of female rats. Brain Res Mol Brain Res 1998; 55: 169–172.
36. D’Souza DN, Zhang Y, Damjanoska KJ, et al. Estrogen reduces serotonin-1A receptor-mediated oxytocin release and Galpha (i/o/z) proteins in the hypothalamus of ovariectomized rats. Neuroendocrinol 2004; 80: 31–41.
37. Birzniece V, Johansson IM, Wang MD, et al. Serotonin 5-HT(1A) receptor mRNA expression in dorsal hippocampus and raphe nuclei after gonadal hormone manipulation in female rats. Neuroendocrinol 2001; 74: 135–142.
38. Lu NZ and Bethea CL. Ovarian steroid regulation of 5-HT1A receptor binding and G protein activation in female monkeys. Neuropsychopharmacol 2002; 27: 12–24.
39. Hay DL and Poyner DR. Calcitonin gene-related peptide, adrenomedullin and flushing. Maturitas 2009; 64: 104–108.
40. Osterlund MK, Halldin C and Hurd YL. Effects of chronic 17β-estradiol treatment on the serotonin 5-HT1A receptor mRNA and binding levels in the rat brain. Synapse 2000; 35: 39–44.
41. Amin Z, Canli T and Epperson CN. Effect of estrogen–serotonin interactions on mood and cognition. Behav Cogn Neurosci Rev 2005; 4: 43–58.
42. Puri V, Cui L, Liverman CS, et al. Ovarian steroids regulate neuropeptides in the trigeminal ganglion. Neuropeptides 2005; 40: 35–46.
43. Schreihofer DA, Rowe DF, Rissman EF, et al. Estrogen receptor-alpha (ERalpha), but not ERbeta, modulates estrogen stimulation of the ERalpha-truncated variant, TERP-1. Endocrinol 2002; 143: 4196–4202.
44. Shughrue PJ, Bushnell CD and Dorsa DM. Estrogen receptor messenger ribonucleic acid in female rat brain during the estrous cycle: a comparison with ovariectomized females and intact males. Endocrinol 1992; 131: 381–388.
45. Yang Y, Ozawa H, Kawata M, et al. Immunocytochemical analysis of sex differences in calcitonin gene-related peptide in the rat dorsal root ganglion, with special reference to estrogen and its receptor. Brain Res 1998; 791: 35–42.
46. Wyon Y, Frisk J, Lundeberg T, et al. Postmenopausal women with vasomotor symptoms have increased urinary excretion of calcitonin gene-related peptide. Maturitas 1998; 30: 289–294.
47. Goadsby PJ, Edvinsson L and Ekman R. Vasoactive peptide release in the extracerebral circulation of humans during migraine headache. Ann Neurol 1990; 28: 183–7.
48. Juhasz G, Zsombok T, Jakab B, et al. Sumatriptan causes parallel decrease in plasma calcitonin gene-related peptide (CGRP) concentration and migraine headache during nitroglycerin induced migraine attack. Cephalalgia 2005; 25: 179–83.
49. Levy D, Burstein R, Kainz V, et al. Mast cell degranulation activates a pain pathway underlying migraine headache. Pain 2007; 130: 166–76.
50. Sarajari S and Oblinger MM. Estrogen effects on pain sensitivity and neuropeptide expression in rat sensory neurons. Experimental Neurol 2010; 224: 163–169.
51. Ma QL, Zhou HY and Sun M. Relationship between sex hormone levels and blood calcitonin gene-related peptide/endothelin-1 in postmenopausal women with coronary heart disease. Hunan Yi Ke Da Xue Xue Bao 2001; 26: 146–148.
52. Corominas R, Sobrido MJ, Ribasés M, et al. Association study of the serotoninergic system in migraine in the Spanish population. Am J Med Genet B Neuropsychiatr Genet 2010; 153B(1): 177–84.
53. Ishii M, Shimizu S, Sakairi Y, et al. MAOA, MTHFR, and TNF-β genes polymorphisms and personality traits in the pathogenesis of migraine. Mol Cell Biochem 2012; 363(1–2): 357–66.