Annals of Neurosciences, Volume 19, Issue 4 (October), 2012
New cellular and molecular approaches to ageing brain
ABSTRACT
The last decade has witnessed a mammoth progress in the area of brain ageing. Recent gene profiling and brain imaging techniques have made it possible to explore the dark areas of ageing neurons in a new molecular perspective. Many conserved pathways and cellular and molecular mechanisms particularly nuclear mitochondrial molecular interactions are known now. Disruptions in mitochondrial function and reduction in cellular antioxidative and immunoproteins contribute to generation of reactive oxygen species (ROS) which leads to deteriorated adult neurogenesis, reduced white matter and compromised neural plasticity. The overall deteriorated structure and function of neurons is manifested in form of cognitive decline and prolonged neurodegenerative disorders. Dietary restrictions (DR), physical and mental activities however have been shown to counter these ailments. However more precise molecular dynamics at protein levels is still debatable which is the future task for neuroscientists.
Keywords: Dietary restriction, ROS, NTFs, Synaptic plasticity, Dentate gyrus, Antioxidant.
Corresponding Author: Anurag Tripathi, PhD, Ranchi University, Email: rahulzau@rediffmail.com, Tel: +91-9955489044.
doi : 10.5214/ans.0972.7531.190410
Introduction
Brain ageing is characterized by many physical, chemical or biological changes in the status of neurons which is often manifested as deterioration in the Cognitive function and demetia.1 This phenominan is one of the most striking because it is the major risk factor for most common neurodegenerative diseases including alzheimer’s disease (AD), parkinson’s disease (PD), amylotrophic lateral sclerosis (ALS) and stroke. Recent studies indicate that normal brain aging is associated with subtle morphological and functional alterations in specific neuronal circuits rather then large scale loss of neurons.2 The loss of neurons, however in normal brain aging is compensated by expanding dendritic arbors and synaptic contacts whilst in age related neurodegenerative disorders, dendritic arbors and synaptic connections are lost and compensation doest not occur.3
In fact, aging of the brain in diverse mammalian species shares many common features such as dendritic regression in pyramidal neurons, synaptic atrophy, decrease of striatal dopamine receptors, accumulation of fluorescent pigments, cyto skeletal abnormalities and reactive astrocytes and microglia.4
Although age associated defects in particular neuronal circuits have been described, the molecular basis of aging brain still remains debatable. Fortunately the last 15 years have witnessed a significant increase in our knowledge of the basic molecular mechanisms of aging. Most remarkably, functional genetic analysis has identified signaling pathways that act as master regulators of ageing and life span that are conserved in yeast, nematodes, flies and mammals.1
However, two important technical advances have provided new insight in to the biology of brain aging. Micro-array technology has made global gene expression analysis possible in human and model organism leading to the identification of evolutionary conserved changes during ageing. Functional brain imaging technology has enabled us to study the cognitive networks in the ageing human brain. The present review seeks to present a discussion on Neuro-anatomical and molecular alterations integrated with ageing process, mitochondrial dysfunction and autophagy related to ageing, alteration in gene expression during normal and degenerative ageing and its interaction with reactive oxygen species. The impact of dietary restriction as a preventive measure has also been discussed.
Neuro anatomical and Cellular Changes during Normal and Degenerative Brain Ageing
Brain aging is characterized by a plethora of anatomical changes which are the consequence of molecular and cellular alterations. Early studies suggested that substantial neuronal loss occurs in the ageing neocortex and hippocampus which are the most vulnerable regions of ageing. However, recent studies showed that neuronal loss was not significant in most regions of the ageing neocortex and hippocampus.5 In contrast dendritic branching could increase in some hippocampal regions in aged individuals6 while ageing prefrontal cortex (PFT) showed variable changes in dendritic branching patterns.5 Many investigations have reported increased dendritic extent in dentate gyrus (a subregion of hippocampus) of old compared with middle aged humans.7 In other subregions of human hippocampus including areas CA18 (Carnuammonis) and CA39 and the subiculum,10 there is no change in dendritic branching with age. The morphology of PFC neurons seems to be more vulnerable to the effects of ageing than that of hippocampal neurons. In rat, dendritic branching of pyramidal neurons decreases with age in superficial Cortical layers.11 A reduction in dendritic branching with age has also been observed in anterior cingulate layer V of rat12 and the human medial PFC13. The data available, however, on spine density also suggested its region specific alterations5. The hippocampal region in aged human showed no significant reduction in spine density.15,16 In addition to these changes reduction in synapse number is also markable in aged brains. An early electron microscopic study at the perforant path granule cell synapse showed 27% decrease in axodendritic synapse number in the middle molecular layer of dentate gyrus in aged rates as compared with that of young rats.17,18 These neuro anatomical changes, however, result in impaired neuro plasticity and ultimately alter the network dynamics of neural ensembles that support cognition. At the cellular level, however, an extensive loss of myelinated nerve fibres from the white matter of the human cerebral hemispheres during normal ageing has been reported19 with the greatest reduction in the PFC and corpus callosum.20,21 In addition, alterations in their myelin sheaths with age are also reported.22 Electron microscopic study has shown that integrity of myelin sheath is disrupted with age. Overall break down of myelin sheaths would cause disruption of conduction along nerve fibres and a reduction in the connectivity between parts of the brain,22 consequently causing reduced speed of information.
Chemical and Neurophysiological Changes
Brain ageing is not merely accompanied by morphological and anatomical deterioration but a large number of neurochemical and neurophysiological alterations are also witnessed in an integrated form. Reductions are found in neurochemical systems most notably in dopaminergic, noradrenergic, and cholinergic pathways23–26 thus resulting in increased cognitive impairments and dementia. In the human and rhesus macaque pre frontal cortex (PFC) the secretion of inhibitory neurotransmitter gama – amino butyric acid (GABA) is diminished in aging brain due to reduced gene expression thus altering the balance between inhibitory and excitatory neurotransmission.27 This may contribute to increased neural activity in PFC which could predispose individuals to excitotoxicity and neurodegenerative pathology. Positron Emission Tomography (PET) in humans have shown significant decrease in dopamine synthesis,28 notably in the striatum and extrastreatal regions excluding mid brain.29 Significant age related decrease in dopamine receptors D1, D2 and D3 are also noticed, particularly receptors binding in to caudate nucleus and putamen.30,31
PET studies in humans have also shown decrease in the level of serotonin receptor S2 in the caudate nucleus, putamen and frontal cortex in aging brains34 as well as a decreased binding capacity of the serotonin transportor, 5HHT, in the thalamus and the mid brain.35 In addition glutamate also shows decreased level in aging brains particularly in parietal gray matter, basal ganglia and frontal white matter.36,37 However, electrical properties of the neurons remain constant over the life span in all the subrigions of the hippocampus.38
In contrast numerous studies have shown an increase in Ca++ conductance in aged neurons. CA1 pyramidal cells in the aged hippocampus have an increased density of L – type Ca++ channels.39 In addition to changes in Ca++ channels, impaired intraneuronal calcium buffering capacity may increase cytoplasmic free Ca++ levels.19
A major neuronal calcium buffering protein, calbindin 1 has been reported to be reduced in basal forebrain cholinergic and cortical neurons in aging human and non-human primates.40 More over these decrements could be attributed to reduced in RNA expressions of calbindin and Ca++ channel genes, in PFC41. It is proposed therefore that impaired Ca++ homeostasis could lead to altered synaptic plasticity.
Alterations in gene expression in ageing brain
There are ample evidence suggesting that cognitive impairments and neurodegenerative disorders may be associated with specific changes in gene expression. Gene expression profiling studies of ageing mouse, rat, monkey and human have shown significant alteration in the expression of synaptic genes.42–45 More than 150 genes have been noted to undergo age-dependent expression changes in these organisms which may be up regulated or down regulated.1
Most of the micro-array studies have shown reduced expression of genes involved in mitochondrial energy metabolism which may become more pronounced in humans with cognitive decline and AD.46–48 Another significant set of genes which shows increased expression during aging is that involved in stress response pathways.1 Gene expression studies of the ageing neocortex in mice, monkey and humans has shown age-dependent up regulation of the apolipoprotein D gene.49 The expression of this gene in Drosophila extends life span playing role as a lipid antioxidant conferring resistance to oxidative stress.49–51 Moreover apolipoprotein D expression is induced in the brains of individuals with AD. In over all picture however genes responsible for glial activity, myelin proteins, metal ion homeostasis, immune response and stress response in humans show upregulated gene expression while genes pertaining to mitochondrial function, neural plasticity, ubiquitin – proteaosome pathways show down regulated expression in human ageing brains.7
In addition, genes involved in synaptic functions that mediate memory and learning including glutamate receptor sub unites, synaptic vesicle proteins and members of major signal transduction systems that mediate long term potentiation showed down regulated expression.19
Moreover, genes involved in stess response including antioxidant defense, DNA repair and immune function constitute largest category of age upregulated genes.19 Among the gene expression profiling studies of the ageing brain to neurodegenerative disorders such as AD, up regulated expression of signaling and tumor suppressor genes and down regulated expression of protein folding, metabolism and energy related gene has been reported.46
Mitochondrial dysfunction
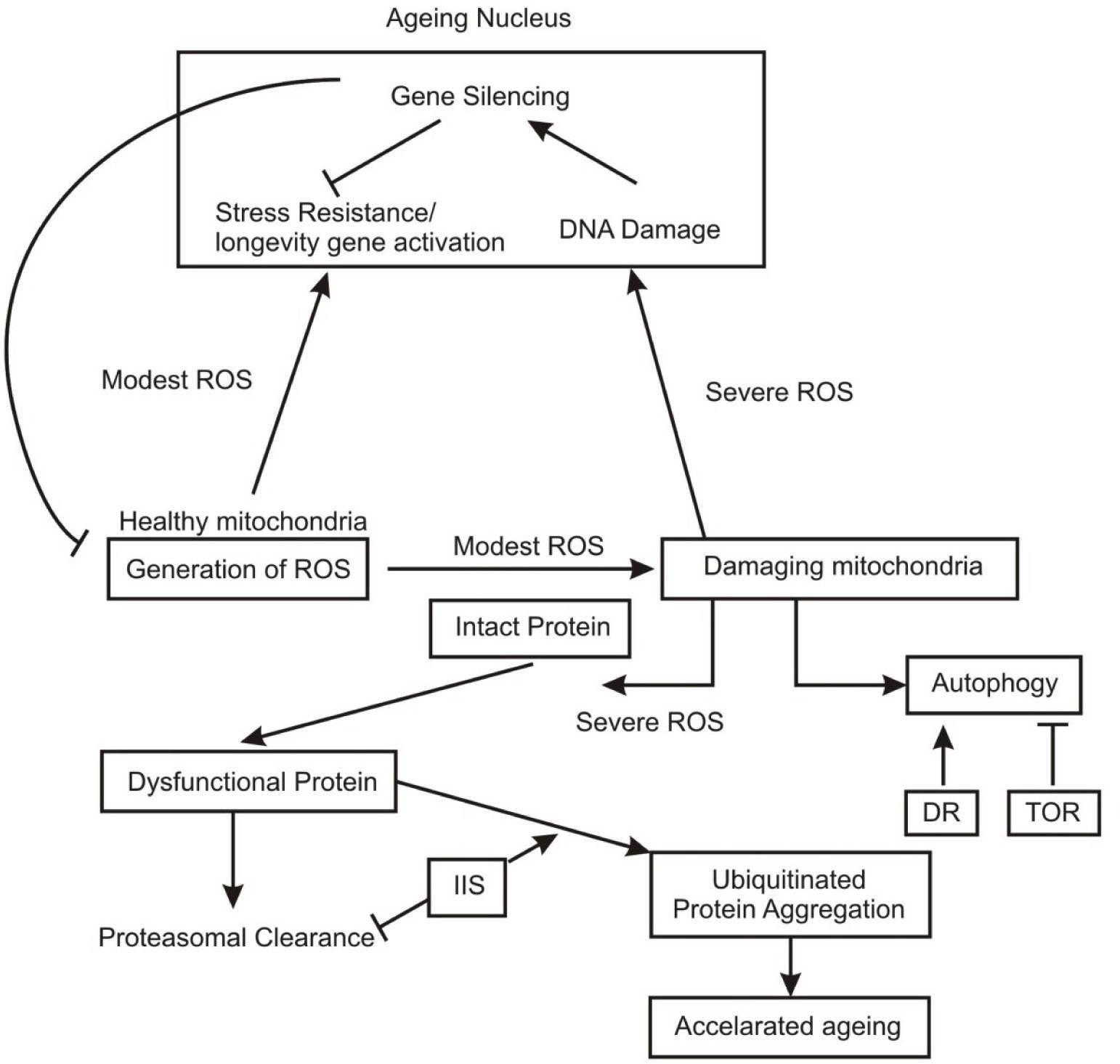
Many gene expression profiling studies have clearly shown a progressive degeneration in mitochondrial function which could contribute to the accelerated ageing particularly in brain, since brain and muscle are more susceptible to mitochondrial dysfunction. Mitochondrial oxidative phophorylation is the key source of energy intensive ion fluxes and axonal transport in the projection neurons of cerebral cortex which degenerate in alzheimer’s desease.19 These neurons, therefore, are highly vulnerable to mitochondrial dysfunction. Respiratory chain enzymes and mitochondrial DNA are the prime targets of mitochondrial damage52 (Fig. 1).
Generation of Reactive Oxygen Species (ROS)
Due to irregularities in the electron transport chain in mitochondria during progressive aging many super oxides are generated as a byproduct which may cause damage to respiratory chain proteins and mitochondrial DNA. In normal course mitochondria passes sufficient machinery to counter these ROS in form of antioxidant enzymes including Cu-Zn super oxide dismutase, cytochrome oxidase and redox reactions mediated by cytochrome C.19 In case of aging, the action of these antioxidants is diminished, resulting in local oxidative damage to mitochondrial proteins and DNA. Super Oxide Dismutase (SOD) reacts with superoxide radicles and converts them in to hydrogen peroxide (H2O2) which is a stable molecule and may diffuse into cytoplasm where it is enzymatically neutralized by cytoplasmic glutathione peroxidase and peroxisomal catalase. However besides super oxides and H2O2, redox mediated iron is a major sourse of ROS mediated cellular damage.19 Elevated levels of redox – active iron accumulates in normal ageing brain and in several neurodegenerative diseases.50
Gene profiling studies, however, have shown the age related reduced expression of mitochondrial genes in organisms ranging from nematode to humans.5,6,19,54,55 Conversely augmented mitochondrial function has been shown to extend life span.56 Targeted over expression of the antioxidant enzyme catalase specifically in rat mitochondria extends the life span.57 Although the actual mechanism that extends the life span in organisms is still debatable, one hypothesis is that efficient electron transport chain (ETC) function reduces the generation and release of ROS. In addition, many nuclear transcripts that declined in the aging brains are required for mitochondria function.54 These included NADP transhydrogenase, ubiquinol – cytochrome C reductase complex, subumite VIII of cytochrome Coxidase and gama and delta subunites of F1 particle.19 All these components are the integral members of ETC. This profile suggests that mitochondria function may be compromised in ageing brains.
Fig. 1: Molecular pathways showing Role of ROS, DR and IIS in neural ageing: Dysfunctional mitochondria and ubiquitinated protein aggregates, promote ageing (—| indicate inhibition;→ shows activation).
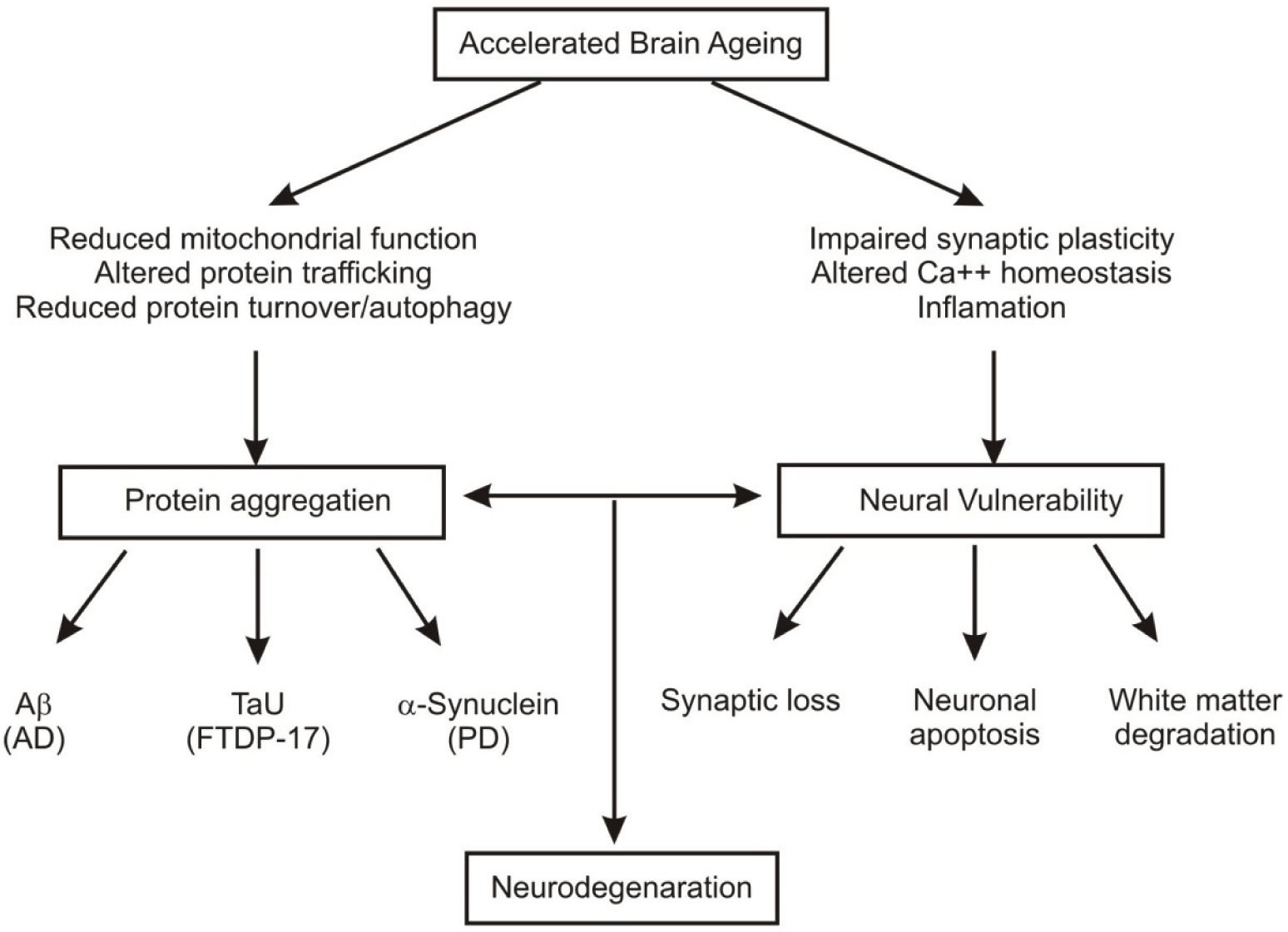
Fig. 2: Neurodegeneration and accelerated brain ageing.
Of late many interesting microarray studies have shown contradictory findings to above mentioned facts regarding role of mitochondrial dysfunction in ageing.1
CKL – I is required for synthesis of ubiquinone, a key component of ETC. CLK – I mutants have reduced respiratory chain but have long life spans.58 Subsequent studies based on RNA interference screens found that reduction of function in many genes affecting ETC can increase life span.59,60 This effect appears to be dose dependent because a modeost reduction in ETC activity can increase life span whereas a more severe reduction shorten it.56 Recent evidence suggests that this life span extension may be mediated by nuclear transcriptional response to mitochondrial defects termed as retrograde response involving the induction of oxidative stress resistance and xenobiotic detoxification genes.61 Moreover in Drosophila and a mouse model with a reduced expression of ETC components in neurons life span is extended.62,63 Intriguingly this mouse model also shows protection against neuronal excitotoxicity.19 The signaling mechanisms, however, mediating increased longevity in this context are not well known. It is probable that ROS in a modestly increased concentration may act as signaling molecule to promote longevity. This observation further raises the possibility that the initial decline in mitochondrial gene expression during brain aging may be a part of an active compensatory mechanism that increases stress resistance.
Autophagy and protein homeostasis as a regulatory mechanisms in aging brains
Recent studies in worms, flies and mouse have established autophagy of mitochondria as a key component to extend life span64–66 and reduced autophagy may contribute to neuro degeneration.66,67 Reduced autophagy, however in neuro degenerating brains in flies and mice is accompanied by aggregation of ubiquitinated proteins, similar to those observed in human neurodegenerative disorder such as Huntington’s disease (HD) and AD.1 The clearance of a mutant protein huntingtin in HD is directly under the regulation of expressed BECN I.68 In addition to BECN I a number of other genes related to autophagy have been reported to be declined in brain ageing (T Lu and BA Yankner, unpublished results). Thus, accumulation of dysfunctional protein in reduced autophagy may contribute to severe ROS generation and the release of redox active iron leading to neuronal impairements.
Two significant pathways accelerating neural aging
Target of rapamycin (TOR) pathway normally inhibits autophagy, contributing in impaired protein homeostasis. Reduced TOR signaling has been reported to extend life span in yeast, worms, flies, mice.69,70 Though the extent to which TOR signaling affects life span is unknown but together with autophagy this has significant role in age dependent neurodegenerative diseases caused by protein aggregates. Another significant signaling mechanism contributing to brain aging is insulin/IGF – I signaling (IIS) pathway. Reduced IIS pathway has been shown to extend life span in worms, flies and mammal.71 In contrast, in mammals, insulin and IGF – I are neurotrophic and promote neuronal survival by inhibiting apoptosis.72 These can also promote learning and memory in humans and animal models.71,72 There exists a dichotomy therefore, between neuro-protective effects of insulin and IGF – I and their adverse effects on life span. Interestingly the effects on life span parallel the effects on neurodegeneration. Knockout mouse of Irs2 or IGF – I receptor can reduce cognitive impairment and neurodegeneration in models of AD.73,74 In patients with AD, reduced expression of IGF signaling is reported.75 The role of IIS pathway is therefore debatable about its response as an effective neuro-protector as well as indicator of neuro-degenerative process.
Delaying the Effects of Neural Ageing
After having a thorough review over cellular and molecular components and pathways of brain ageing, it is worthwhile to discuss the preventive measures of cognitive impairments and neuro degenerative processes. Brain supportive healthy diets including omega 3 fatty acid, vitamin C, vitamin E (an effective anti-oxidant) vitamin B12, vitamin B6, folic acid iron, calcium, zink, docosa hexaenoic acid (DHA) and breast milk proteins have been primarily reported to delay the effects of normal brain aging and cognitive decline.
Recent investigation on the impact of dietary restriction (DR) as brain aging and neuro degenerative disorders have shown many striking features. Dietary restriction (reduction in diet without causing malnutrition) has been reported to play multi-dimentional roles at cellular and molecular levels. DR has been reported to reduce age related gene expression alterations upto substantial level associated with stress and immune responses respectively.54 These effects of DR on immune stress related transcripts indicates that both autoimmunity and oxidative damages are reduced in the brains of DR mice.76 In addition to its suppressive role on many gene transcripts, DR is shown to induce many gene expressions.
One of the largest classes of transcripts induced by DR (9%) comprised growth and neurotrophic factors including the develop mentally regulated homeobox genes which might be involved in neural development and gene encoding neuroserpin, a factor that promotes neural plasticity.3 Other transcripts that are induced by DR include transforming growth factor (TF) and brain derived neurotrophic factor (BDNF) which can protect neurons against excitotoxic and metabolic insults.77 Other genes to be induced under DR influence are that related to DNA synthesis. This observation might be related to increased neurogenesis in rodents under DR.78 These gene profiling studies are supportive of the fact that modulation of energy metabolenin, oxidative stress, ion homeostasis by DR could affect brain ageing in the mouse3 (Fig. 3). More recent studies have shown that enhancement of BDNF and other neurotrophic factors due to DR, exerts beneficial effects on synaptic plasticity and might therefore facilitate learning and memory.79 The capacity of the brain for neurogenesis might decrease in ageing80 and DR has been shown to increase the number of newly generated neural cells in dentate gyrus of the rat hippocampus.78 HSP – 70 and GRP – 78 proteins which protect neurons against excitotoxic and oxidative insults81,82 have been noted to be increased in cortical, strial and hippocampal neurons of DR rats.83,84
In addition to its protective role in normal brain ageing, DR is reported to exert protective role against neurodegenerative disorder AD85 and PD.86
However in addition to multifaceted contribution of DR in protection of normal brain ageing and neurodegenerative disorder, regular physical exercise has been shown to increase neurogenesis and neurotrophic factors.87 Over the past decade a number of epidemiological studies have shown a decreased risk for PD and dementia of subjects who exercise regularly.88–90
Conclusion
Gene expression profiling and brain imaging techniques have given a new insight to the cellular, molecular and behavioral alterations in aging brains. After having an overview over the recent developments on neural ageing, in the present review neural ageing has been discussed as the consequence of decreased neurogenesis and synaptic plasticity, altered neuro chemical and signaling pathways, reduced white matter, mitochondrial dysfunction, enhanced stress responses and accumulation of ROS and dysfunctional proteins, reduced antioxidative, DNA repair enzymes and decreased immune responses etc. At the molecular level, however, it is due to alterations in the expression of a wide array of genes involved in these processes.
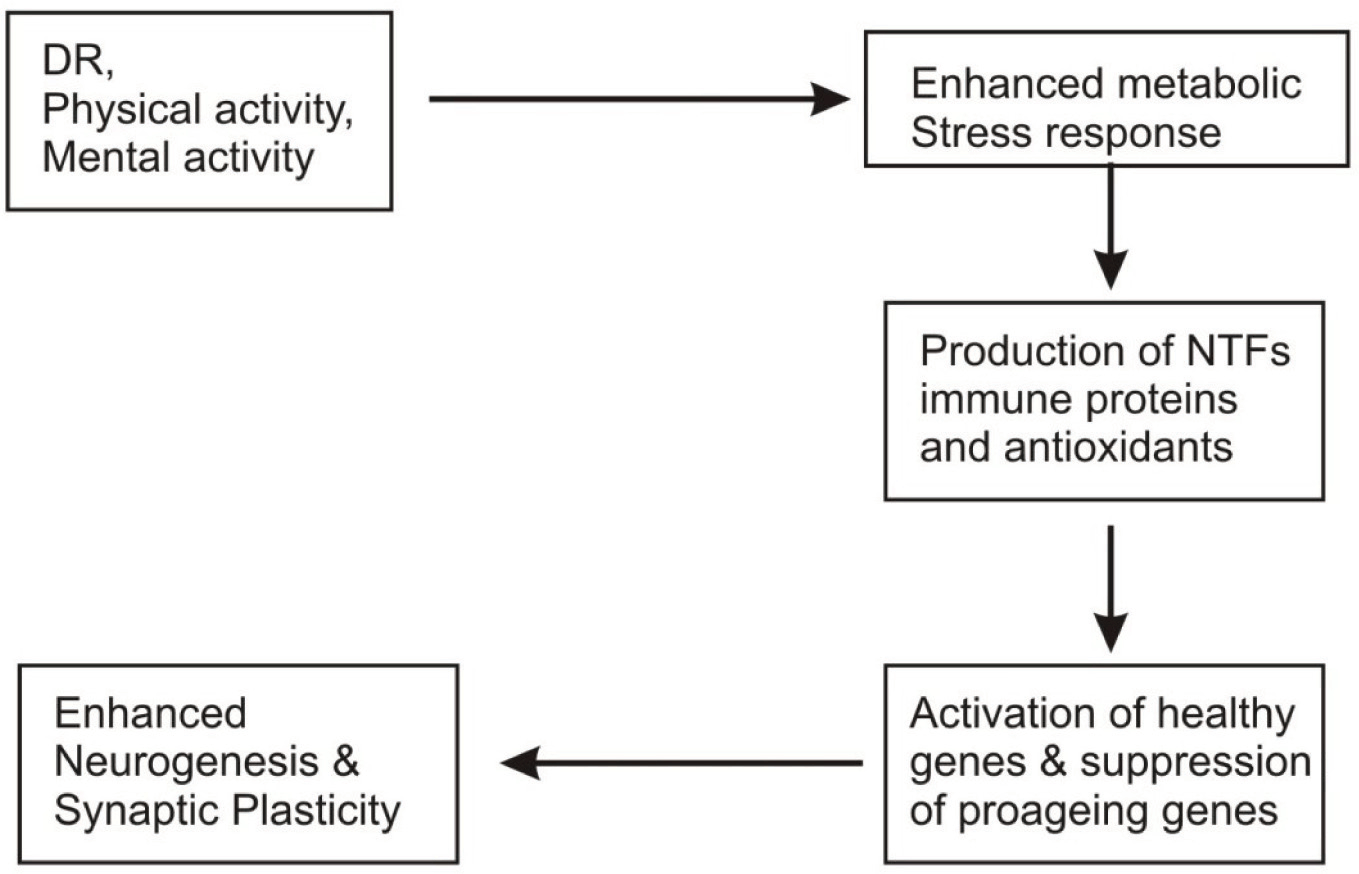
Fig. 3: Anti ageing components and their role in prevention of brain ageing.
Moreover, neocortex, hippocampus and striatal centres are the most vulnerable areas, affected in aging with a variable degree of changes in their subcentres. Therapeutic and preventive measures have also been briefly discussed, with particular reference to DR since recent studies are focused on it. Hence, management of these canditions through medical and life style interventions is likely to benefit in order to cope with these age related impairments. Moreover, the function of nervous system depends upon highly specific intricate intercellular signaling networks whose regulatory mechanisms extend beyond gene transcription. It is, therefore essential to understand such mechanisms at the level of protein interactions within individual cells, organelles and synapses.3 It is, therefore, imperative prospect to explore this molecular dynamics through combined proteomic and brain imaging techniques in a more comprehensive manner.
Acknowledgement
The present research work is supported by UGC — MRP No. F PSJ - 007/10: 11, sanctioned to the author.
Abbreviations used in Figs. 1–3
| ROS | = | Reactive Oxygen species |
| DR | = | Dietary Restriction |
| TOR | = | Target of Rapamycin |
| IIS | = | Insulin Growth Factor - 1 signalling |
| AB | = | β amyloid |
| AD | = | Alzheimer›s disease |
| PD | = | Parkinsen›s disease | NTF | = | Neurotrophic factor |
| FTDP-17 | = | Frontotemporal dementia with parkinsonsims linked to chromosome - 17 |
The article complies with International Committee of Medical Journal Editor’s uniform requirements for the manuscripts.
Competing interests: None, Source of funding: UGC
Received Date : 10 May 2012
Revised Date : 01 July 2012
Accepted Date : 30 September 2012
References
1. Bishop NA, Lu T, Yanker BA. Neural mechanisms of ageing and cognitive decline. Nature 2010; 46: 529–535.
2. Morrison JH and Hof PR. Life and death of neurons in the ageing brain. Science, 1997; 278: 412–419.
3. Prolla TA and Mattson MP. Molecular mechanisms of brain ageing and neurodegenerative disorders: lessions from dietary restriction. TINS 2001; 24(11): 521–530.
4. Wanagat J. Caloric intake and ageing: mechanisms in rodents and a study in nonhuman primates. Taxicol Sci. 1999; 52: 35–40.
5. Burke SN and Barnes CA. Neural plasticity in the ageing brain. Nat. Rev. Neurosci. 2006; 7: 30–40.
6. Buell SJ and Coleman PD. Dandritic growth in the aged human brain and failure of growth in senile dementia. Science 1979; 206: 854–856.
7. Flood DG, Buell SJ, Horwitz GJ, etal. Dendritic extent in human dentate gyrus granule cells in normal ageing and senile dementia. Brain Res. 1987; 402: 205–216.
8. Hanks SD and Flood DG. Region specific stability of dendritic extent in normal human ageing and regression in alzheimer’s disease. I.CA1 of Hippocampus. Brain Res. 1991; 540: 63–82.
9. Flood DG, Guarnaccia M and Caleman PD. Dandritic extent in human CA2–3 hippocampal pyramidal neurons in normal ageing and senile dementia. Brain Res. 1987; 409: 88–96.
10. Flood DG. Region specific stability of dendritic extent in normal human ageing and regression in alzheimer’s disease. II subiculum. Brain Res. 1991; 540: 83–95.
11. Grill JD and Riddle DR. Age related and laminar specific dendritic changes in the medial frontal cortex of the rat. Brain Res 2002; 937: 8–21.
12. Markham JA and Juraksa JM. Ageing and sex influence the anatomy of the rat anterior cingulate cortex. Neurobiol. Ageing 2002; 23: 579–588.
13. De Brabander JM, Kramers RJ and Uylings HB. Layer specific dendritic regression of pyramidal cells with ageing in the human pre frontal cortex. Eur J. Neurosci. 1998; 10: 1261–1269.
14. Uylings HB and de Brabander JM. Neuronal changes in normal human ageing and alzheimer’s disease. Brain Cong. 2002; 49: 268–276.
15. Williams RS and Metthysse S. Age related changes in Down Syndrom brain and the cellular pathology of alzheimer’s disease. Prog. Brain Res. 1986; 70: 49–67.
16. Curio CA and Hinds JW. Stability of synaptic density and spine volume in dentate gyrus of aged rats. Neurobiol. Ageing 1983; 4: 77–87.
17. Bondareff W and Geinisman Y. Loss of neurons in the dentate gyrus of the senescent rat. Am J Anat 1976; 145: 129–136.
18. Gemisman Y, Bodareff W and Dodge JT. Partial deafferentation of neurons in the dentate gyrus of the senescent rats. Brain Res. 1977; 134: 541–545.
19. Yankner BA, Lu T and Loerch P. The ageing bran. Annu. Rev. Patho. Mech. Dis 2008; 3: 41–66.
20. Heden T and Gabrieli JD. Insights into the ageing mind: a view from cognitive neuroscience. Nat. Rev. Neurosci. 2004; 5: 87–96.
21. Bartzokis G, Cummings JL, Sultzer D, et al. White matter structural integrity in healthy ageing adults and patients with Alzheimer disease: A magnetic resonance imaging study. Arch Neurol. 2003; 60: 393–938.
22. Peters A and Rosene DL. In ageing is it gray or white? J Comp Neurol 2003; 462: 139–143.
23. Eriksdotter–Nilsson M, Garhardt G, Granholm AC, et al. Multiple changes in noradrenergic mechanisms in the coeruleo– hippocampal pathway during ageing. Structural and functional correlates in intraocular double grafts Neurobiol ageing 1989; 10: 117–124.
24. Jouvenceau A, Dutar P, Bellard JM. Alterations of NMDA receptor-mediated synaptic responses in CA1 area of the aged rat hippocampus: Contribution of GABA ergic and cholinergic deficits. Hippocampus 1998; 8: 627–637.
25. Wese RA. Dopamine learning and motivation. Natl. Rev. Neurosci. 2004; 5: 597–609.
26. Floel A and Cohen LG. Translationl study in neuro-rehabilitation: From bench to beside. Cogn. Behv. Neurol 2006; 19: 1–10.
27. Loerch PM, Jianbin W, Kelly AD et al. Evaluation of the ageing brain transcripteome and synaptic regulation. Plos one. 2008; 3: e 3329.
28. Hof PR and Mobbs CV. Handbook of the neuroscience ageing. 2009; London, UK: Elsevier Inc.
29. Ota M and Yasuno F, Suhara T, et al. Age related decline of dopamine synthesis in the living human brain measured by positron emission tomography with L–Dopa. Life Sciences 2006; 79: 730–736.
30. Kaasinen V, Vilkman H, Hietala J, et al. Age related dopamine D2/D3 repotor loss in extras streatal regions of the human brain. Neurobiol. Ageing. 2000; 21: 683–688.
31. Wang Y, Chan GLY, Schulzer M, et al. Age dependent decline of dopamine D1 receptors in human brain: A PET study. Science 1998; 30: 56–61.
32. Iyo M and Yamasaki T. The detection of age related decrease of dopamine D1 D2 and serotonin S – HT2 receptors in living human brain. Prog. Neuro-psychophermacol and Biol Psy chait. 1993; 17: 415–421.
33. Juha OR, Pirkko L, Schulzer M et al. Age dependent decline in human brain dopamine D1 and D2 receptors. Brain Research 1989; 508: 349–352.
34. Wong DF, Wanger HN, Douglass KH, et al. Effects of age on dopamine and serotonin receptors measured by PET in the living human brain. Science 1984; 266: 1393–1396.
35. Yamamoto M, Suhara T, Yoshiro O, et al. Age related decline of serotonin transporters in living human brain of healthy males. Life Sciences. 2001; 71: 751–757.
36. Chang L, Jiang CS and Ernst T. Effects of age and sex on brain glutamate and other metabolites. MRI 2009; 27: 142–145.
37. Saliasuta N, Chang L and Ernst T. Regional variations and the effects of age and sex on brain glutamate and other metabolites. Magn Reson Imaging 2008; 26: 667–675.
38. Barnes CA. Normal aging: regionally specific changes in hippocampal synaptic transmission. Trends Neurosci 1994; 17: 13–18.
39. Thibault O and Landfield PW. Increase in single L-type calcium channels in hippocampal neurons during aging. Science 1996; 272: 1017–1020.
40. Geula C, BU J, Nagykery N, et al. Loss of calbindin – D 28K from ageing human cholinergic basal fore brain: relation to neuronal loss. J. Comp Neurol 2003; 455: 249–259.
41. Lu T, Pan Y, Kao SY, et al. Gene regulation and DNA damage in the ageing human brain. Nature 2004; 429: 883–891.
42. Jiang CH, Trien JZ, Hu Y, et al. The effects of aging on gene expression in the hypothalamus and cortex of mice. Proc Natl Acad Sci USA 2001; 98: 1930–1934.
43. Blalock EM, Nada MP, Keith S, et al. Gene microarrays in hippocampal ageing: Statistical profiling identifies novel processes correlated with cognitive impairments. J. Neurosci 2003; 23: 3807–3819.
44. Fraser HB, Khaitovich P, Eisen MB, et al. Aging and gene expression in the primate brain. PL0S Biol. 2005; 3: e274.
45. Erraji – Benchekroun L, Arango V, Underwood MD, et al. Molecular ageing in human prefrontal cortex is selective and continuous throughout adult life. Biol Psychiatry, 2005; 57: 549–558.
46. Blalock EM, Geddes JW, James WG, et al. Incipient alzheimer’s disease: Microarray correlation analysis reveal major transcriptional and tumor suppressor responses. Proc. Natl. Acad. Sci. USA, 2004; 101: 2173–2178.
47. Liang WS, Eric MR, Jon V, et al. Alzheimer’s disease is associated with reduced expression of energy metabolism genes in posterior singulate neurons. Proc. Natl. Acad. Sci. USA, 2008; 105: 4441–4464.
48. Miller JA, Oldham MC, Geschwind DH. A system level analysis of transcriptional changes in alzheimer diseases and normal aging. J Neuosci. 2008; 28: 1410–1420.
49. Walker DW, Muffat J, Benzer S, et al. Over expression of a Drosophila homolog of apolipopretein D leads to increased stress resistance and extended life span. Curr Biol. 2006; 16: 674–679.
50. Sanchez D, Begona LA, Laura T, et al. Loss of glial lazarillo, a homolog of apolipoprotein D, reduces life span and stress resistance in Drosophila. Curr. Biol. 2006; 16: 680–686.
51. Kalman J, Mc Conathy W, Kasa P, et al. Apolipoprotein D in the ageing brain and in alzheimer’s dementia. Neurol Res. 2000; 22: 330–336.
52. Wallace DC. A mitochondrial paradigm of metabolic and degenerative diseases, ageing and cancer: A dawn for evolutionary medicine. Annu. Rev Genet 2005; 39: 359–407.
53. Zecca L, Youdim MB, James RC, et al. Iron, Brain aging and neurodegenerative disorders. Nat. Rev Neurosci. 2004; 5: 863–873.
54. Lee CK, Weindruch R, Prolla TA. Gene expression profile of the ageing brain in mice Nature Genet 2000; 25: 294–297.
55. Zahn JM, Art BO, Suresh P, et al. Age map: a gene expression data base for ageing in mice. PLOS. Genet. 2007; 3: e 201.
56. Rea SL Vetura N and Johnson TE. Relationship between mitochondrial ETS dysfunction, development and life extension in Coenorrhabditis elegans. PLOS Biol. 2007; 5: e 259.
57. Schriner SE, Nancy JL, George MM, et al. Extension of murine life span by over expression of catalase targeted to mitochondria. Science 2005; 309: 481–484.
58. Branicky R, Benard C and Hekimi S. Clk – 1 mitochendria and physiological rates. Bio Essays 2000; 22: 48–56.
59. Lee SS, Lee RY, Fraser AG, et al. A systematic RNAi screening idetifies a critical rate for mitochendria in C. elegans longevity. Nature Genet. 2003; 33: 40–48.
60. Dillin A, Hsin H, Fraser AG, et al. Rates of behavior and ageing specified by mitochondrial function during development. Science 2002; 298: 2398–2401.
61. Cristina D, CaryM, Lunce ford A, et al. Regulated response to impaired respiration slows behavioral rates and increases life span in C elegans. PLOS Genet. 2009; 5: e100450.
62. Copeland JM, Hur JH, Bahadorani S, et al. Extension of Drosophila life span by RNAi of the mitochondrial respiratory chain Curr Biol. 2009; 19: 1591–1598.
63. Dell agnello C, Leo S, Agostino A, et al. Increased longevity and refractoriness to Ca++ dependent neurodegeneration in Surf 1 knockout mice. Hum. Mol Genet 2007; 16: 431–444.
64. Melendez A, Tallóczy Z, Seaman M, et al. Autophagy genes are essential for daver development and life span extension in C. elegans. Science 2003; 301: 1387–1391.
65. Simonsen A, Schubert DR, Finley KD, et al. Promoting basal levels of autophagy in the nervous system enhances longevity and oxidant resistance in adult drosophila. Autophagy 2008; 4: 176–184.
66. Hara T, Kenji N, Makoto M, et al. Suppression of basal autophagy in neural cells causes neurodegenerative diseases in mice. Nature 2006; 441: 885–889.
67. Komatull, Satoshi W, Tomoki C, et al. Loss of autophagy in the central nervous system causes neurodegeneration in Mice. Nature 2006; 441: 880–884.
68. Sibata M, Degterev A, Mizushima N, et al. Regulation of intracellular accumulation of Mutant Huntingtin by Beclin 1. J Biol Chem. 2006; 281: 14474–14485.
69. Schieke SM and Finkel T. Mitochondrial signaling TOR and life span. Biol Chem. 2006; 387: 1357–1361.
70. Harrison DE, Anderson RM, Johnson SC, et al. Rapamycin fed late in life extends life span in genetically heterogeneous mice. Nature 2009; 460: 392–395.
71. Broughton S and Partridge L. Insuin/IGF like signaling, the nervous system and ageing. Biochem, 2009; 418: 1–12.
72. Vander Heide LP, Ramakers GMJ and Smidt MP. Insulin signaling in the central nervous system. Learning to survive. Prog. Neurobiol. 2006; 79: 205–221.
73. Freude S, Roth J, Kahn CR, et al. Neuronal IGF – 1 resistance reduces AB accumulation and protects against premature death in a model of alzheimer’s disease. FASEBJ, 2009; 23: 3315–3324.
74. Cohen E, Du D, Estepa G, et al. Reduced IGF – 1 signaling delays age associated proteotoxicity in mice. Cell 2009; 139: 1157–1169.
75. Molony AM, Ravid R, O’Neill C, et al. Defects in IGF – 1 receptor insulin receptor and IRS – ½ in alzheimer’s disease indicate possible resistance to IGF – 1 and insulin signaling. Neurobiol ageing 2008; 31: 224–243.
76. Sohal RS. Effect of age and caloric restriction on DNA oxidative damage in different tissues of C56 BL/6 mice. Mech. Ageing Dev. 1994; 76: 215–224.
77. Mattson M and Lind Vall O. Neurotrophic factors and the aging brain. (Mattson MP and Geddes JW eds) 1997; pp. 299–345 JAI Press.
78. Lee J, Duan W, Long JM, et al. Dietary restriction increases survival of newly generated neural cells and induces BDNF expression in the dentate gyrus of rats. JMol Neurosci. 2000; 15: 99–108.
79. Black IB. Trophic regulation of synaptic plastisity. J Neurobiol, 1999; 4: 108–118.
80. Kuhn HG, Dickinson-Anson H, Gage FH. Neurogenesis in the dentate gyrus of the adult rat: age related decrease of neuronal progenitor proliferation. J Neurosci 1996; 16: 227–2033.
81. Lowenstein DH, Chan PH, Miles MF. The stress protein response in cultured neurons: Characterization and evidence or a protective role in excito-toxicity Neuron 19; 7: 1053–1060.
82. Yu ZF, Fu W, Mattson MP, et al. The endoplasmic reticulum stress responsive protein GRP 78 protects neurons against excitotoxcity and apoptosis: suppression of oxidative stress and stabilization of calcium homeostasis; Exp Neurol 1999; 155: 302–314.
83. Duan W and Mattson MP. Dietary restriction and Z-deoxyglucose administration improve behavioural outcome and reduce degeneration of dopaminergic neurons in models of Parkinson’s disease. J Neuroci Res 1999; 57: 195–206.
84. Yu ZF and Mattson MP. Dietary restriction and Z-deoxyglucose administration reduce focal ischemic brain demage and improved behavioral outcome: evidence or a preconditioning mechanism. J Neurosci Res. 1999; 57: 830–839.
85. Mayux R. Reduce risk of alzheimer’s disease among individuals with low coloric intake. Neurology 1999; 59(Suppl): 296–297.
86. Logroscino G, Tang MX, Shea S, et al. Dietary lipids and anti-oxidants in parkinson’s disease: A population based, case control study. Ann Neurol 1996; 39: 89–94.
87. McCarter RJ. Role of caloric restriction in the prolongation of life. Clin Geriatr. Med. 1995; 11: 553–565.
88. Sohal RS and Weindruch R. Oxidative stress, Caloric restriction and ageing. Science 1996; 273: 59–63.
89. Frame L, Hart RW, and Leakey JE. Caloric restriction as a mechanism mediating resistance to environmental disease. Environ Health Perspect 1998; 106 (Suppl): 313–324.
90. Masoro EJ. Influence of caloric intake on ageing and on the response to stressors. J Toxicol Environ Health 1998; 1: 243–257.