Annals of Neurosciences, Volume 20, Issue 2 (April), 2013
Spatially regulated adult neurogenesis
ABSTRACT |
Adult neurogenesis has been the center of attention for decades. Neuroscientists hope to understand the mechanism underlying this phenomenon that might provide a unique perception of brain repair in future. Neurogenesis is referred to the process in which neuronal stem cells and progenitors generate new neurons in non-pathologic setting. Although there are some similarities between two neurogenetic regions including hippocampus and olfactory bulb, however there are some important differences. Regardless of the unique functional roles of ongoing neurogenesis in olfactory bulb and hippocampus, the differences are in terms of consequence of neurogenesis, origin of newly born neurons, responding receptors to nicotine exposure, neuronal migration and GABAergic input between two regions. In this paper, we have briefly reviewed the differences of adult neurogenesis between olfactory bulb and hippocampus. KEYWORDS: Olfactory bulb, Hippocampus, Central nervous system, Neuronal migration, GABAergic signalling Corresponding Author: Nusrat J. Mobassarah, Ph.D., Institute of Integrated Cell-Material Science, Kyoto University, Yoshida Ushinomiyacho, Sakyo – ku, Japan, 606-8501, Tel.: 81-80-757539869, Fax: 81-80-757539820, E-mail: snusratjahan@icems.kyoto-u.ac.jp doi : 10.5214/ans.0972.7531.200208 |
Introduction
The century old dogma of neurobiology was based on the doctrine stated by the prominent histologist Ramon Y Cajal that “In the adult centres, the nerve paths are something fixed, ended and immutable. Everything may die, nothing may be regenerated”.1,2 According to this prevailing hypothesis, the process of regeneration of functional neurons from progenitor cells could only occur in embryonic state of human brain.1,2 However, contrary to this old dogma, past decade witnessed tremendous novel studies in neuroscience yielding to the acceptance of the phenomena of adult neurogenesis.2-9 Several studies have demonstrated that in adult mammalians brain neurons undergo mitosis, migrate, and integrate into existing circuitries.2-9 The process of adult neurogenesis is also shown to exist in two specific regions such as dentate gyrus hippocampus and subventricular zone of the lateral ventricle or olfactory bulb, which are referred as “neurogenic regions”.6-9 All stages of neuronal development do occur during adult neurogenesis as it occurs as part of embryonic and postnatal development processes such as mitosis of precursor cells and formation of mature and functioning new neurons. Although there are some similarities between neurogenesis in olfactory bulb and hippocampus, there are distinct differences in certain aspects of neurogenesis in these two neurogenic regions. In this article, we have analysed such spatially regulated differences in the process of neurogenesis.
Major differences between two neurogenic brain regions
Neuronal replacement versus neuronal addition
Imayoshi and his colleagues studied labeled neuron stem cells (NSCs) in R26-LacZ mice in which tamoxifen(antagonist of estrogen receptor in breast tissue) was administered for 4 days at the age of 2 months.10 Careful analysis of labeled cells showed increase proliferation of these cells in granular layer of olfactory bulb. In addition, peri-glomerular cells were also found to have increased. After 6 months, labeled granular cells reached to 40% and increased to about 50-60% after 12-18 months. Stability in the number of labeled cells could be due to labeling saturation. This observation also indicated that although small population of pre-existing cells remained the same without replacement, but highly significant numbers of pre-existing cells in olfactory bulb were replaced with new labeled neurons by 12 months of tamoxifen treatment. According to these findings, it could be concluded that neurogenesis is an important factor contributing to neuronal replacement in olfactory bulb throughout the life.10
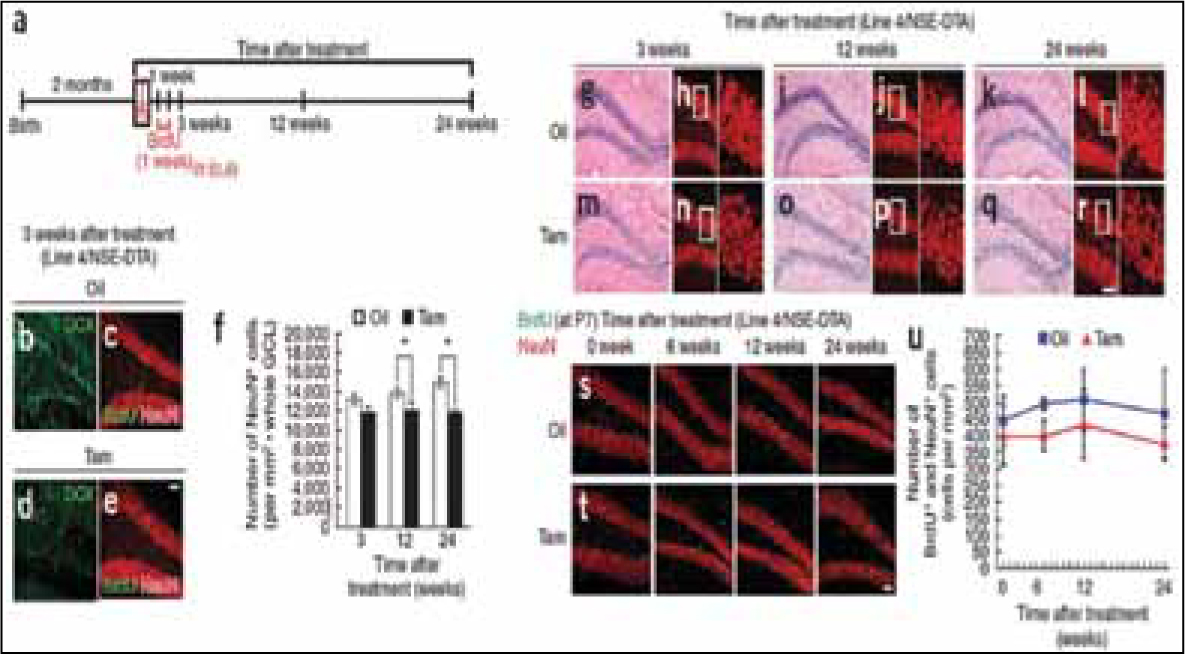
The same research group also studied R26-CFP mice, administered tamoxifen at 2-month age, but examined labeled cells in dentate gyrus.10 Unlike in olfactory bulb, newly born labeled granular cells increased and integrated into granular layers in the dentate gyrus. The increase in number also corresponded to density of granular cells. This study also showed that ablation of neurogenesis in mice hindered increase in number of granular cells in dentate gyrus without any effect on neuronal circuitries (Fig. 1). This study clearly highlighted the distinct consequence of neurogenesis in dentate gyrus and olfactory bulb. In the case of dentate gyrus, neurogenesis results in increased number of neuronal cells whereas in olfactory bulb pre-existing neurons gets replaced since the size of olfactory bulb is fairly stable throughout lifetime.10
Neuronal migration
In adult mice, though distance between subventricular zone (SVZ) and olfactory bulb is only a few millimetres, but for a very small neuron cell it is not easy to travel. Neuronal cells initially migrate along the wall of the lateral ventricle then along rostral migratory stream (RMS) to the olfactory bulb. The cells move together and form a chain along RMS (Fig. 2).11,12 Contrary to cell migration in olfactory bulb; newly born cells in dentate gyrus take a short journey to reach inner granular cells. Rapid extension of axons of immature granular cells into CA3 area of hippocampus has been observed during four to eight days after mitosis (Fig. 3).13
Fig. 1: Ablation of neurogenesis inhibits the increase of the granule cell number in the dentate gyrus. (a) Experimental design. (b–e) The number of newly formed granule cells (DCX+ and BrdU+NeuN+) decreased in the dentate gyrus of tamoxifen-treated Line 4/NSE-DTA mice compared with the oiltreated mice.(f) Quantification of the number of NeuN+ granule cells in the dentate gyrus of oil- and tamoxifen-treated Line 4/NSE-DTA mice. (g–r) HE staining (g,i,k,m,o,q) and immunostaining for NeuN (h,j,l,n,p,r) of the dentate gyrus of oil- and tamoxifen-treated Line 4/NSE-DTA mice. (s,t) Early-born NeuN+ granule cells labeled with BrdU survived similarly in oil- and tamoxifen-treated Line 4/NSE-DTA mice. (u) Quantification of the number of NeuN+BrdU+ granule cells in the dentate gyrus of oil- and tamoxifen-treated Line 4/NSE-DTA mice. NeuN+ granule cells were labeled with BrdU before oil or tamoxifen treatment. *P 0.05, t test. Scale bars represent 50 mm. Nat Neurosci. 2008;11:1153–61.
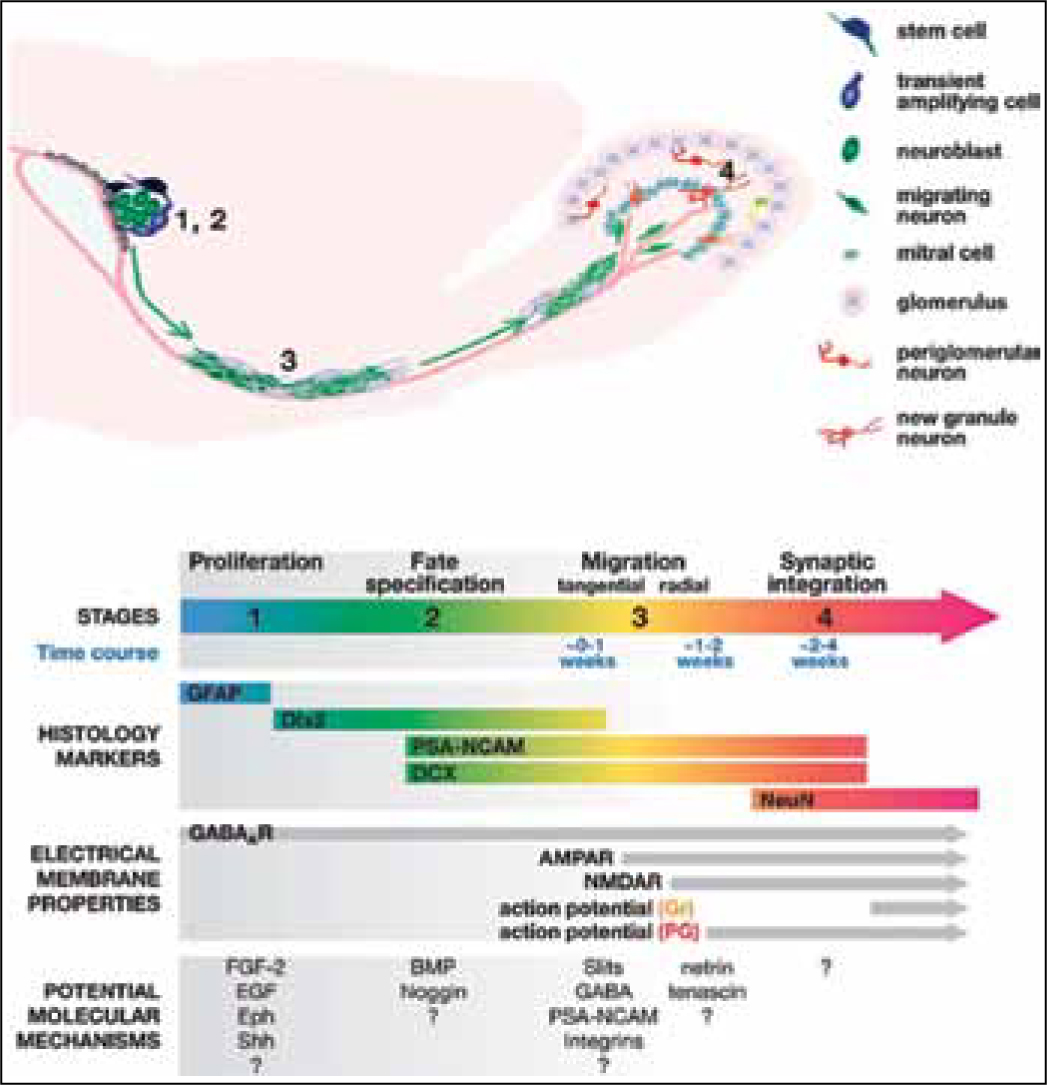
Fig. 2: Generation of new interneurons in the olfactory bulb from neural stem cells in the subventricular zone (SVZ). Adult neurogenesis in the SVZ/olfactory systems undergoes four developmental stages. Stage 1.Proliferation: stem cells (blue) in the SVZ of the lateral ventricles give rise to transient amplifying cells (light blue). Stage 2. Fate specification: transient amplifying cells differentiate into immature neurons (green). Adjacent ependymal cells (gray) of the lateral ventricle are essential for neuronal fate specification by providing inhibitors of gliogenesis. Stage 3. Migration: Immature neurons (green) migrate with each other in chains through the rostral migratory stream (RMS) to the olfactory bulb. The migrating neurons are ensheathed by astrocytes. Once reaching the bulb, new neurons then migrate radially to the outer cell layers. Stage 4. Synaptic integration: Immature neurons differentiate into either granule neurons (Gr, orange) or periglomerular neurons (PG, red ). These unusual interneurons lack an axon and instead release their neurotransmitter from the dendritic spines at specialized reciprocal synapses to dendrites of mitral or tufted cells. The specific properties of each stage are summarized below, mainly on the basis of studies in adult mice. Annul. Rev. Neurosci. 2005. 28:223–50.
Responding receptors to chronic nicotine exposure
Mechawar et al investigated the effect of chronic exposure to nicotine on mice.14 Twenty-one days after labeling cells with 5-bromodeoxyuridine (5’BrdU), number of labeled cells in granular cell layer in olfactory bulb of mice chronically exposed to nicotine was much lower than the same cells in olfactory bulb of mice exposed to saline (Fig. 4).14 Presence of the same number of labelled cells in nicotine and saline exposed mice lacking β2-nAChR, a high-affinity receptor for nicotine, confirmed critical role of these receptors in apoptosis. It can be concluded that these types of nicotinic receptors are actively involved in survival of new born granular cells in olfactory bulb.14
Study done by Arbrous and his colleagues showed that nicotine self-administration also decreased the number of Brdu-IR in granular cell layer of dentate gyrus of male Sprague dawley rats in a dose dependent fashion.15 Unlike in olfactory bulb, effect of nicotine is not mediated by activation β2-nAChR in dentate gyrus; instead nicotinic receptor called alpha-bungarotoxin (alpha-Bgt-AChRs) induces suppressor protein 53 and the cdk inhibitor p21. Since nicotine is cytotoxic to undifferentiated cells in hippocampus, alpha-Bgt-AChRs receptors participate in neuronal proliferation not differentiation in hippocampus.16 The role of alpha-Bgt-AChRs in hippocampus differs from the role that β2-nAChR plays in survival of newly born neurons in olfactory bulb.
Sources of newly born neurons in neurogenic regions
Previous studies have showed that multi-potential and self-renewing stem cells in subependyma are sources for newly born neurons traveling along RMS and join to olfactory bulb as interneurons.17 It was assumed that stem cells residing in sub granular layer of adult hippocampus are source of newly born neurons. One study tested this assumption by using two different methods, which yielded same results. The study indicated that although stem cells with long-term self-renewing and multipotentiality reside in adjacent-subependyma regions, but neuronal stem cells do not exist in dentate gyrus. The study also suggested neuronal progenitors with limited self-renewal capacity as the source of newly generated neurons in adult dentate gyrus of rodent.17
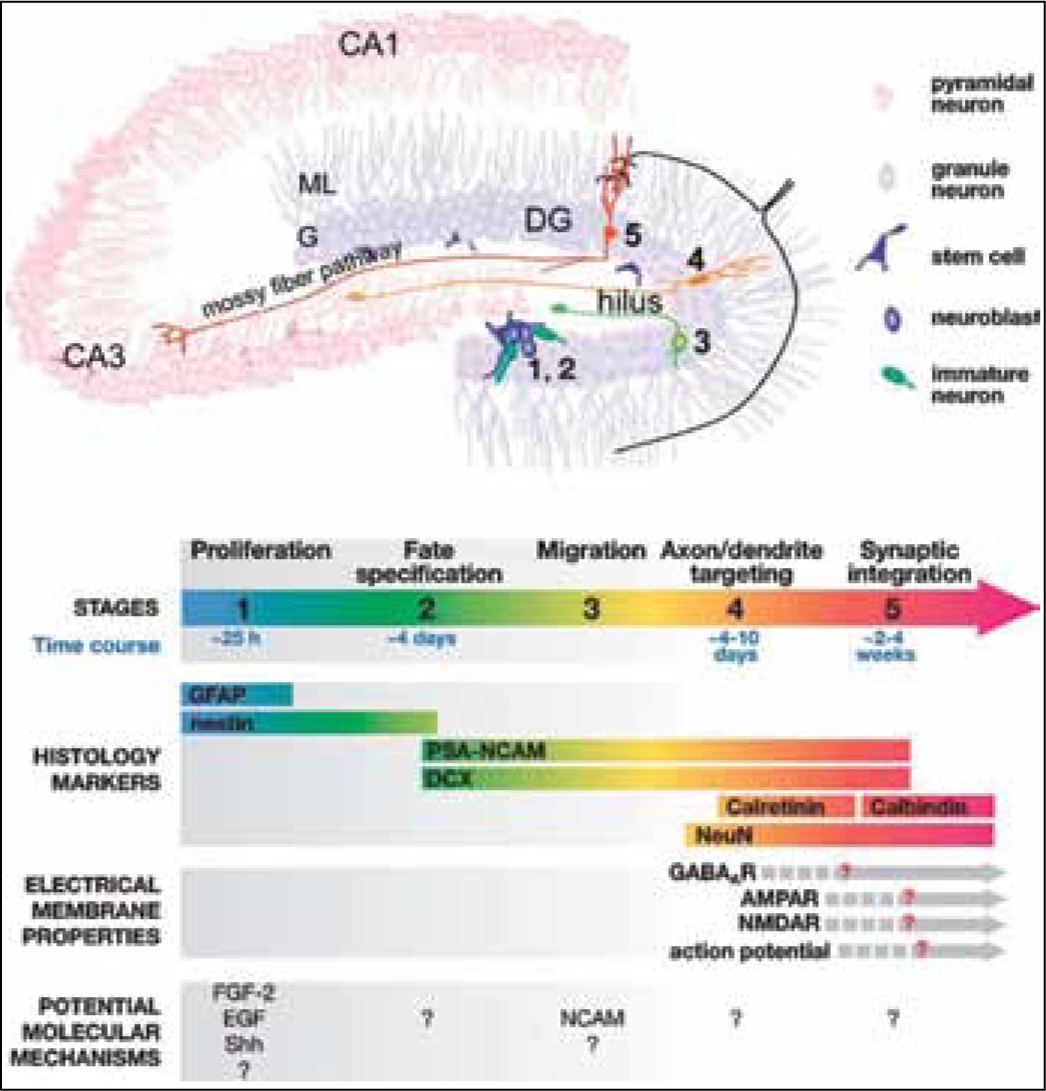
Fig. 3: Generation of new granular neurons in the dentate gyrus of the hippocampus from neural stem cells in the subgranular zone (SGZ). Adult neurogenesis in the dentate gyrus of the hippocampus undergoes five developmental stages. Stage 1. Proliferation: Stem cells (blue) with their cell bodies located within the subgranular zone in the dentate gyrus have radial processes that project through the granular cell layer and short tangential processes that extend along the border of the granule cell layer and hilus. These stem cells give rise to transient amplifying cells (light blue). Stage 2. Differentiation: transient amplifying cells differentiate into immature neurons (green). Proliferating progenitors in the SGZ are tightly associated with astrocytes and vascular structures. Stage 3. Migration: Immature neurons (light green) migrate a short distance into the granule cell layer. Stage 4. Axon/dendrite targeting: Immature neurons (orange) extend their axonal projections along mossy fiber pathways to the CA3 pyramidal cell layer. They send their dendrites in the opposite direction toward the molecular layer. Stage 5. Synaptic integration: New granule neurons (red) receive inputs from the entorhinal cortex and send outputs to the CA3 and hilus regions. The specific properties of each stage are summarized below, mainly on the basis of studies in adult mice. DG, dentate gyrus region; ML, molecular cell layer; GL, granular cell layer. Neurosci. 2005. 28:223–50.
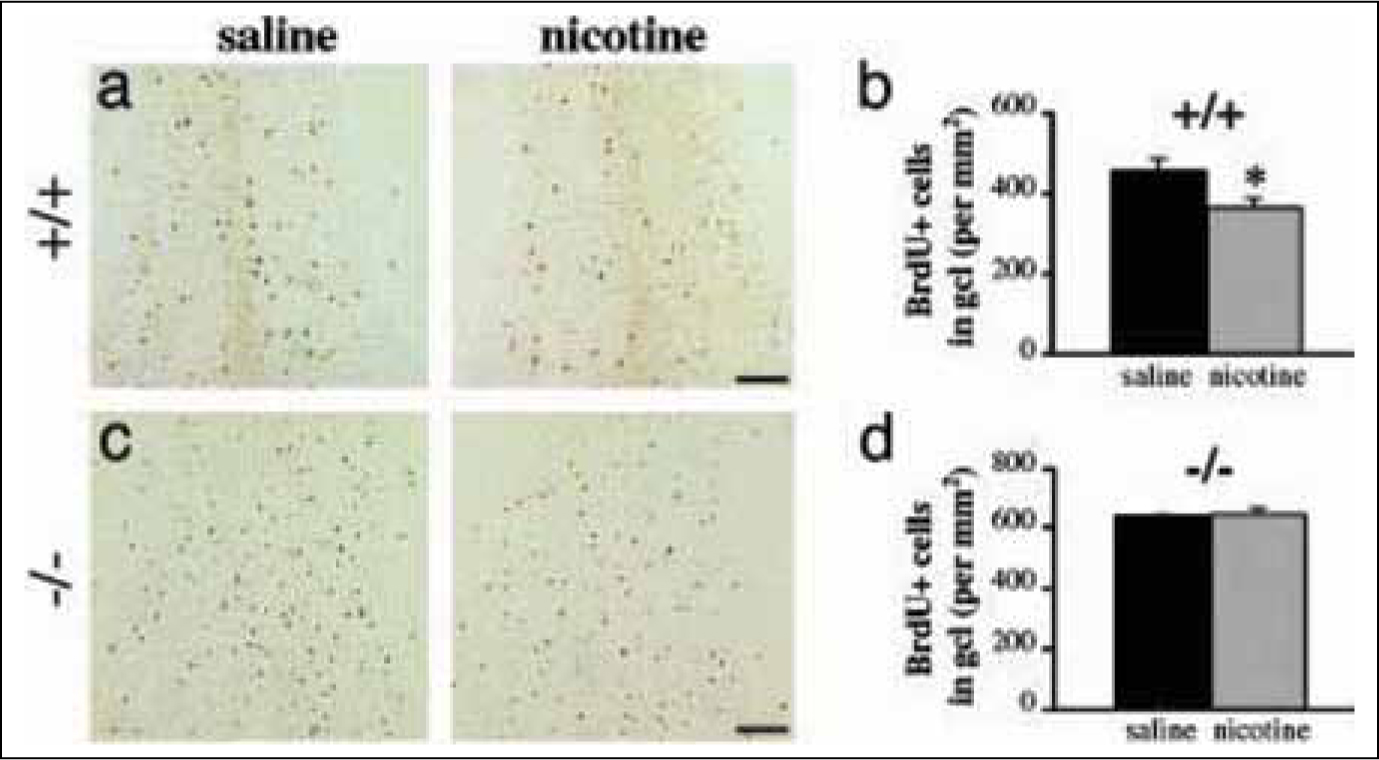
Fig. 4: Chronic nicotine exposure decreases survival of newborn OB neurons via *β2-nAChR activation. (a and c) Representative micrographs of BrdUrd+ cells in the granule cell layer of chronic saline- and nicotine-exposed β2+/+ and β2-/- mice, and corresponding densities of BrdUrd+ cells (b and d). β2+/+, n = 6 per group; β2-/-, n = 3 per group. *, P<0.05 by Student’s t test. (Scale bars: 100 μm.) Proc Natl Acad Sci U S A. 2004 June 29;101(26):9822–9826.
Non-synaptic versus synaptic release of GABA
Liu et al. demonstrated the existence of non-synaptic GABAergic signalling between neuroblasts and glial fibrillary acidic protein (GFAP).18 In this study, inhibitation of GABA receptors increased the number of GFAP-cell expressing proliferation whereas stimulation of GABA release from neuroblasts of SVZ stem cell progeny attenuated GFAP-cell proliferation.18
In another interesting study, it is showed that type-II cells that express GABA receptors and actively divide into progenitors cells in adult dentate gyrus receive GABAergic input from local mature hippocampal circuitry. It is also logical to suggest that neuronal progenitor cells in SGZ have synapses with GABAergic interneurons in hippocampus.19 According to the result of these two studies, non-synaptically and synaptically released GABA affect proliferation of stem cells in SVZ and type-II cells in dentate gyrus of hippocampus.18,19
Conclusion
Deeper understanding of differences of adult neurogenesis in olfactory bulb and hippocampus at molecular and cellular level could be a prerequisite for use of neuronal stem cells for therapies in future. The knowledge of different mechanisms involving conducting proliferation, differentiation, and apoptosis of neuronal stem cells in olfactory bulb and hippocampus not only enables selection of appropriate stem cells based upon the patient’s need but also addresses the issue of personalized treatment of brain injury or brain disease by individualizing the cell culture environment.
The article complies with International Committee of Medical Journal Editor’s uniform requirements for the manuscripts.
Competing interests – None, Source of Funding – None
Received Date : 3 January, 2013
Revised Date : 27 February, 2013
Accepted Date : 25 March, 2013
References
1. Ramon y Cajal S. Degeneration and regeneration of the nervous system. London: Oxford Univ. Press 1913.
2. Colucci-D’ Amato L , Bonavita V, di Porzio U. The end of the central dogma of neurobiology: stem cells and neurogenesis in adult CNS. Neurol Sci 2006; 27: 266–270.
3. Gage FH. Neurogenesis in the adult brain. J. of Neurosci. 2002; 22(3): 612–613.
4. Altman J, Das GD Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J Camp Neurol 1965; 124: 319–335.
5. Sahay A, Scobie KN, Hill AS, et al. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 2011; 472: 466–472.
6. Deng W, Aimone JB, Gage FH. New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nature Rev Neurosci 2010; 11: 339–350.
7. Ming G, Song H. Adult neurogenesis in the mammalian brain : significant answers and significant questions. Neuron 2011; 70: 687–702.
8. Doetsch F, García-Verdugo JM, Alvarez-Buylla A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J Neurosci 1997; 17: 5046–5061.
9. Garcia-Verdugo JM, Ferron S, Flames N, et al. The proliferative ventricular zone in adult vertebrates: A comparative study using reptiles, birds and mammals. Brain Res Bull 2002; 57: 765–775.
10. Imayoshi I, Sakamoto M, Ohtsuka T, et al. Roles of continuous neurogenesis in the structural and functional integrity of the adult forebrain. Nat Neurosci 2008; 11: 1153–1161.
11. Lois C, García-Verdugo JM, Alvarez-Buylla A. Chain migration of neuronal precursors. Science. 1996; 271: 978–981.
12. Ming G, Song H. Adult neurogenesis in the mammalian central nervous sytem. Annu. Rev. Neurosci 2005; 28: 223–250.
13. Hastings NB, Gould E. Rapid extension of axons into the CA3 region by adult-generated granule cells. J Comp Neurol 1999; 11: 297–320.
14. Mechawar N, Saghatelyan A, Gould E, et al. Nicotinic receptors regulate the survival of newborn neurons in the adult olfactory bulb. Proc Natl Acad Sci USA 2004; 101: 9822–9826.
15. Abrous DN, Adriani W, Montaron MF, et al. Nicotine self-administration impairs hippocampal plasticity. J Neurosci 2002; 22: 3656–3662.
16. Berger F, Gage FH, Vijayaraghavan S. Nicotinic receptor-induced apoptotic cell death of hippocampal progenitor cells. J Neurosci 1998; 18: 6871–6881.
17. Seaberg R.M, van der Kooy D. Adult rodent neurogenic regions: the ventricular subependyma contains neural stem cells, but the dentate gyrus contains restricted progenitors. J Neurosci 2002; 22: 1784–1793.
18. Liu X, Wang Q, Haydar TF, et al. Nonsynaptic GABA signalling in postnatal subventricular zone controls proliferation of GFAP-expressing progenitors. Nat Neurosci 2005; 8: 1179–1187.
19. Tozuka Y, Fukuda S, Namba T, et al. GABAergic excitation promotes neuronal differentiation in adult hippocampal progenitor cells. Neuron. 2005; 47: 803–815.