Annals of Neurosciences, Volume 21, Issue 2 (April), 2014
Olfactory Sensory Neuron Morphotypes in the Featherback Fish, Notopterus notopterus (Osteoglossiformes: Notopteridae)
KEY WORDS |
ABSTRACT |
|
Olfactory sensory neurons |
Background: As in other vertebrates, olfactory sensory neurons (OSNs) in fishes are the main components of sensory part of olfactory epithelium that relay olfactory information (smell and taste) to the brain. Purpose: Objective of the present study was to analyze if any polymorphism occurs in the OSNs in a featherback fish, Notopterus notopterus as far as the teleost lineage is concerned. Methods: With the help of neuronal staining technique, polymorphism of OSNs in N. notopterus was studied. Results: Three polymorphic forms of OSNs were identified which are ciliated OSNs, microvillus OSNs and crypt OSNs. These morphotypes were identified on the basis of location of their somata within the depth of olfactory epithelium and resulting length of their dendrites. The ciliated OSNs have basally situated somata and long, thin dendrites with a few apically arranged cilia while microvillous OSNs have somata located midway in the epithelium and thick moderate-length dendrites with microvilli. Third cell type is crypt OSNs which are spherical or pear-shaped, located apically just close to the epithelial surface having cilia and microvilli in an invagination and devoid of any dendrite. Conclusion: N. notopterus belongs to order Osteoglossiformes which is a representative of an early evolutionary lineage of teleost fishes. OSN polymorphism reported in the present work indicates that it is a fairly conserved trait throughout the evolution of teleosts. To our knowledge, we are the first ones to report OSN polymorphism in a member of the order Osteoglossiformes. doi : 10.5214/ans.0972.7531.210205 |
|
Corresponding Author: |
Introduction
Olfactory system is one of the crucial chemosensory systems for fishes functioning in almost all aspects of their life including food-finding, nest-finding, detecting and avoiding predators or other perilous situations,1 feeding,2 kin discrimination,3 reproduction4 and to identify natal streams during spawning migration by salmon.5-7 Fish communication is often accomplished via the use of pheromones.8 In fish and other vertebrates, this system consists of a peripheral part called olfactory rosette (OR) including olfactory nerve formed by the axons of olfactory sensory neurons (OSNs) and a central part comprising olfactory bulb and higher brain areas involved in processing of olfactory information.9 The olfactory epithelium consists of lamellae whose surfaces include sensory and nonsensory regions. Main components of sensory region are the OSNs.
In teleosts, OSNs exhibit three polymorphic forms - ciliated OSNs, microvillous OSNs and crypt receptor cells.10-19 These morphotypes are distinguished by location of their somata within the depth of olfactory epithelium and resulting length of their dendrites. These polymorphs differ in relation to their shape and position within the OE and also functionally.20 They have differential projections to the olfactory bulb,18,19,21,22 different molecular23,24 and physiological properties.18,25-28
Among the teleosts, OSN polymorphism has been reported in many species including the members of cypriniformes - goldfish, Carassius auratus;29 zebrafish, Danio rerio;13 Aplochelius lineatus and Xiphophorus helleri,30 siluriformes - channel catfish, Ictalurus punctatus29,31 and salmonid fishes.16,32
In the present study using the Kluver and Barrera (1953)33 neuronal staining technique, we sought to analyze OSN polymorphism in the olfactory epithelium of a freshwater teleost, featherback fish, Notopterus notopterus belonging to the order Osteoglossiformes.
Methods
Teleost phylogeny
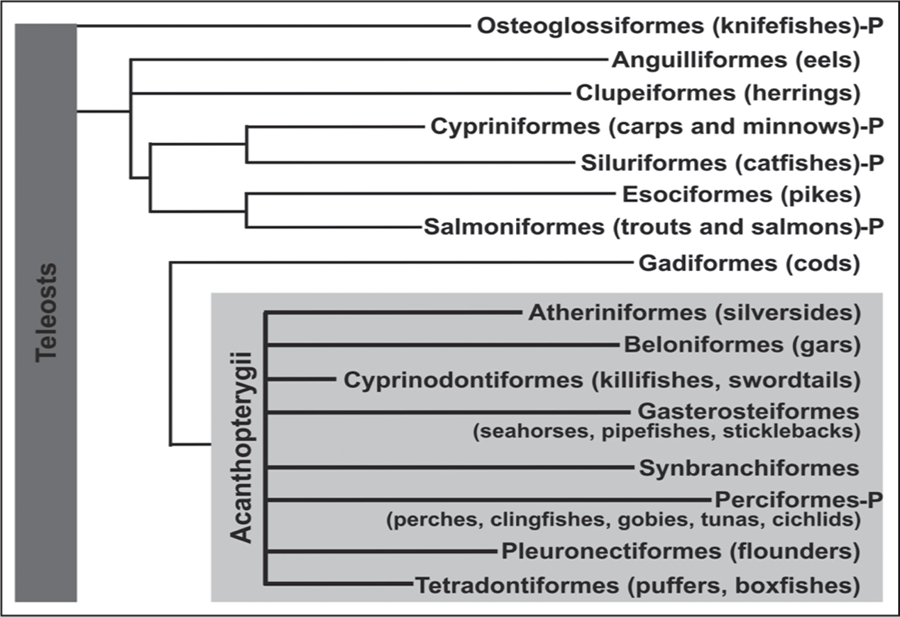
Our study on olfactory sensory neuron morphotypes makes use of fish taxonomy by Nelson (1994)34 and the phylogenetic tree of evolution of teleosts is presented in Fig. 1.
Fig. 1: Phyletic tree of the Teleostei, based on Nelson (1994) showing the occurrence of olfactory sensory neurons polymorphism (P) (For references see the discussion section). The grey box/overlay depicts the group of Acanthopterygii.
Animals
Adult featherbacks (N. notopterus) of either sex (n = 6) with body weight ranging between 125g to 150g and length 27±2cm were obtained from a single freshwater body, Telangkhedi-Futala lake, Nagpur City from 2009-2011. After transport, fishes were maintained in well-aerated glass aquaria (3×2×1.5). Animal care protocols were approved by the Institutional Animal Care and Use Committee.
Neuroanatomical analysis
Fishes were anaesthetized with an aqueous solution of 2 phenoxyethanol (0.03%; P 1126; Sigma), decapitated, olfactory system with the brain was dissected out and immediately fixed in aqueous Bouin’s fixative for 24 hrs, dehydrated in graded series of alcohol and embedded in paraffin wax after clearing in xylene. For neuroanatomical studies, sections of olfactory epithelium were cut at 10 µm thickness in horizontal as well as saggital planes on a rocking microtome, mounted on Mayer’s albumin coated slides, and then subjected to Kluver and Barrera (1953)33 staining. The stained sections were analyzed on a Nikon Eclipse E200 photomicroscope (Japan) and different cellular groups were identified according to their characteristic size, shape, staining intensity of the perikaryon and packing density as well as distribution pattern of the cell bodies. Cell and nuclear diameter were measured with an oculometer. All the numeral data in the results were presented as mean values ± standard deviations (SD).
Analysis of OSN morphotypes
OSN morphology was classified based on depth of soma within the olfactory epithelium and resulting length of their dendrites.20,26 To determine the depth, olfactory epithelium was divided into 3 arbitrary horizontal layers, most apical layer being layer 1 and the most basal being layer 3 (Fig. 3C). With these criteria, OSNs were grouped into one of the three overall types: ciliated, microvillous and crypt types.
Imaging
Desired fields from various sections were photographed using Nikon (E8400) camera at different magnifications and adjusted for size, contrast and brightness in Adobe Photoshop 7.0 and Corel Photo-Paint X4 software. Photo plates were prepared using Corel Draw X4 (version 14) software. Different types of identified OSNs from the photographs were isolated and presented/illustrated according to their distribution in different zones within the olfactory epithelium (Fig. 3B, C). Scale bars were expressed in terms of µm and measurements were taken by using an oculometer.
Results
In the featherback fish, N. notopterus (Fig. 2A), paired olfactory organs are situated on snout region in a cavity called olfactory pits or olfactory chambers connected to the telencephalic hemispheres of brain by a long olfactory tract and are thus pedunculated (Fig. 2B). Each olfactory organ is a cup shaped elongated structure possessing a series of 74±2 lamellae radiating from a central raphe on both the sides (Fig. 2B, C). The lamellae in the middle of rosette (on both sides) are the largest and they gradually taper towards anterior and posterior ends of the rosette (Fig. 2C). Olfactory epithelium (OE) is a thick sheet (30–35 µm) of pseudo-stratified ciliated epithelial cells which is folded to form olfactory lamellae (Fig. 3A). Each lamella is divisible into sensory and nonsensory regions (Fig. 3A). The sensory region is located at the base of lamellae consisting of bipolar OSNs, supporting cells and basal cells (Fig. 3B, C).
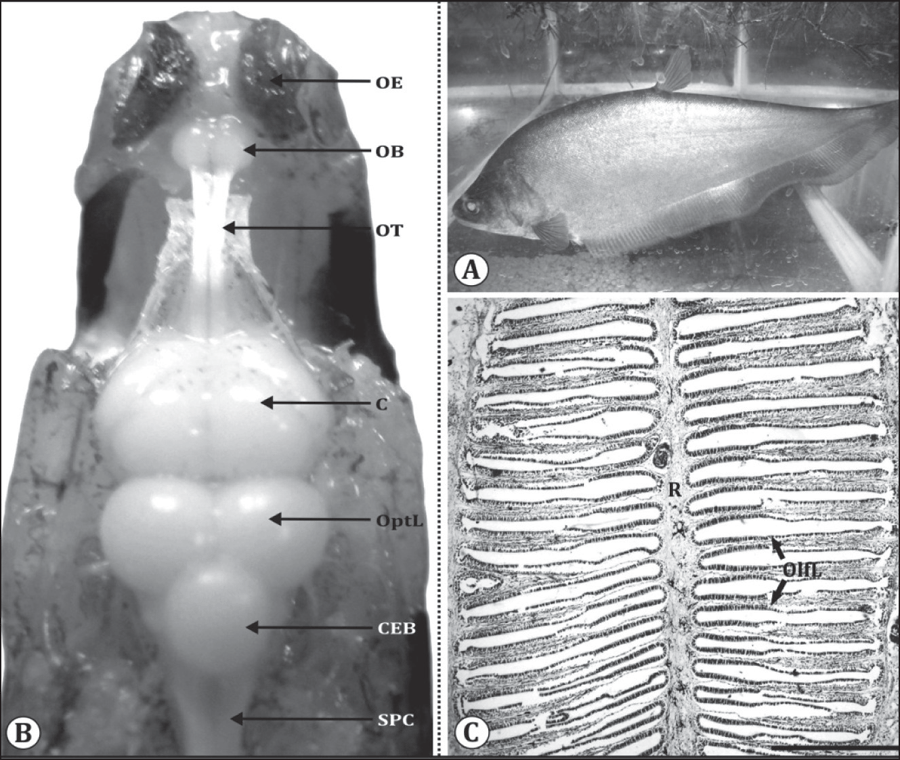
Fig. 2: (A) Photograph of Notopterus notopterus. (B) In situ photograph of olfactory organ with brain of N. notopterus showing; olfactory epithelium (OE), olfactory bulb (OB), olfactory tract (OT), cerebrum (C), optic lobe (OptL), cerebellum (CEB) and spinal cord (SPC). (C) Horizontal section of the olfactory epithelium showing olfactory lamellae (OlfL) radiating from the central raphe (R). Scale bar = 500 μm.
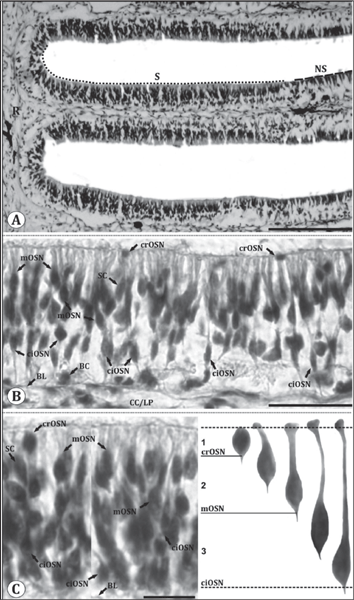
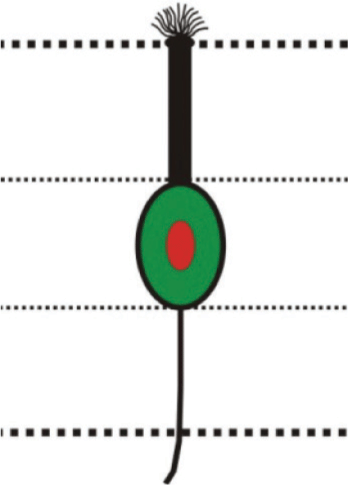
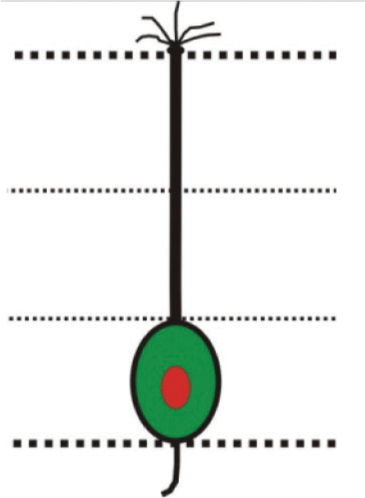
Fig. 3: (A) Part of horizontal section of olfactory epithelium showing location of sensory (S) and nonsensory (NS) regions of olfactory lamellae. Scale bar = 100 μm. (B) Magnified view of sensory region of olfactory lamellae showing; basal cell (BC), basal lamina (BL), central core (CC)/lamina propria (LP), crypt olfactory sensory neurons (crOSNs), ciliated olfactory sensory neurons (ciOSNs), microvillous olfactory sensory neurons (mOSNs) and supporting cell (SC). Scale bar = 50 μm. (C) Magnified view of sensory region of olfactory lamellae (left) showing location of different cell types including; crOSNs, ciOSNs and mOSNs in different zones of olfactory epithelium. Scale bar = 25 μm. Illustrations (right) of different types of olfactory sensory neurons found in different layers of olfactory epithelium (Upper dotted line represents epithelial surface and lower dotted line represents basal lamina).
OSN Morphotypes
In N. notopterus, three types of OSNs are noted- ciliated, microvillous and crypt cells (Fig. 3B, C). Each morphotype is characterized by a cell soma in a specific layer of the olfactory epithelium, variable length of dendrite and extension of axonal process towards basal lamina. OSN morphotypes and their properties are summarized in Table 1.
Table 1: Summary of olfactory sensory neuron morphotypes and their properties.
| OSN Morphotypes | Crypt Cell | Microvillous | Ciliated |
|---|---|---|---|
| Structural organization | 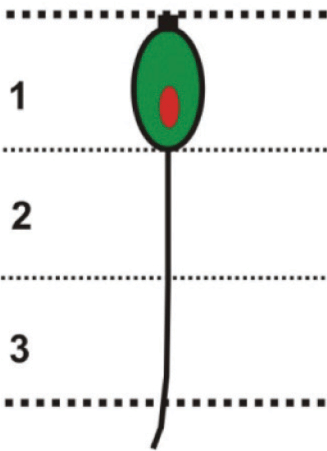 |
 |
 |
| Apical surface | Submerged crypt with cilia and microvilli | Short olfactory knob with microvilli | Olfactory knob with cilia |
| Dendrite | Absent | Thick | Thin |
| Location of cell body in OE | Upper third | Mid region | Lower third |
| Size | 2.9±0.22 μm | 12.06±0.81 μm | 18.78±1.2 μm |
1. Ciliated olfactory sensory neurons (ciOSNs): These are longest (18.78±1.2 µm) among all the types of OSNs distributed throughout the sensory epithelium. These are characterized by their columnar, bipolar organization having basally located somata within layer 3 of the epithelium and their thin, long dendrites reaching upto epithelial surface. They have pronounced olfactory knob with cilia projecting into olfactory mucosa and at the opposite pole axonal process extends towards the basal lamina (Fig. 3B, C). Their cell body containing a round prominent nucleus is situated deep in the epithelium, cytoplasm is highly granular and intensely stained (Fig. 3B, C).
2. Microvillous olfactory sensory neurons (mOSNs): These have moderate (12.06±0.81 µm) length and are characterized by their columnar, fusiform, bipolar organization having cell body located in the mid region within layer 2 of the epithelium. Their thick, moderate length dendrite reaches upto epithelial surface having less pronounced olfactory knob with microvilli at the top. At the opposite pole, axonal process extends towards the basal lamina (Fig. 3B, C). The cell body is more superficial in the epithelium than the ciOSNs and possesses a round nucleus. Cytoplasm is granular and stained intensely (Fig. 3B, C). Population of these cells dominates over the ciOSNs.
3. Crypt olfactory sensory neurons (crOSNs): These are the shortest (2.9±0.22 µm) OSNs. These neurons are characterized by their spherical, pear-shaped or ovoid structure having somata situated apically in the most superficial layer 1 of the olfactory epithelium, devoid of any dendrite and with a axonal process extending toward the basal lamina (Fig. 3B, C). These cells bear submerged cilia and microvilli in the upper portion of the cell and are intensely stained (Fig. 3B, C). They occur regularly in all the lamellae but their absolute number appears to be low as compared to the other two cell types.
Discussion
Olfactory epithelium in N. notopterus is a continuous thick sheet of pseudo-stratified columnar epithelial cells; 30–35 µm in thickness. It is 35 µm in piranha, Serrasalmus nattereri,35 35–55 µm in European eel, Anguilla anguilla36 and 60–75 µm in swordtail, Xiphophorus helleri37 which is folded to form olfactory lamellae. Number, shape and arrangement of lamellae vary considerably among different teleosts ranging from flat unfolded surface to multi-lamellar rosette.9,38-43 Olfactory epithelium in N. notopterus is a multi-lamellar rosette comprising large number (74±2) of lamellae. Generally lamellae are arranged laterally around a central raphe in Zebrafish, Danio rerio,13 Catfish, Clarias batrachus,44,45 Carp, Cirrhinus mrigala,46 Cichlid, Oreochromis mossambicus,47 A. Anguilla,14 Mugil parsia,48 Indian major carp, Labeo rohita,49,50 Wallago attu51 and Macrognathus aculeatus,52 same arrangement is observed in N. notopterus. However, in some other teleosts, olfactory lamellae are arranged at the top of raphe, parallel to each other and in rostro-caudal orientation as in Channa punctatus53 and Channa gachua.54
In N. notopterus, each olfactory lamella comprises sensory and nonsensory regions. Location of these regions varies in different species.42 In N. notopterus, sensory region is at the proximal end and basal region of lamellae and non sensory region at the middle of lamellae. Sensory region is at the middle of lamellae and nonsensory region is at the proximal and basal regions of lamellae in a Cyprinid, L. rohita49,50 but in Rhodeus amarus, sensory region is at the base and middle of lamellae and nonsensory region is at the proximal end.55 Sensory region comprises olfactory sensory neurons, supporting cells and basal cells.13,14 Same cell types are observed in N. notopterus.
OSNs in N. notopterus exhibit polymorphisms similar to that observed in other teleosts. Three OSN morphotypes (ciliated, microvillous and crypt) vary in their shape and position within the epithelium. First type, the ciliated OSN, has a cell body located deep in the OE (in layer 3) near the basal lamina. Its dendrite is long and also quite thin, with a bulbous ending in the olfactory mucosa known as the olfactory knob, which bears cilia.13,14,16,17,20,26,56 Same types of ciOSNs are observed in N. notopterus. Second morphotype, the microvillous OSN, have their cell bodies located in the middle third (in layer 2) of OE having thick and moderately long dendrites bearing microvilli at the top which are also reported in other teleosts.13,14,16,17,20,26,56 These two polymorphic forms are seen even in the rainbow trout embryos (Salmo gairdneri), where ciliated OSNs appear 8 days earlier than the microvillous type.57 Third OSN morph is the crypt cells.11-13 It is devoid of any dendrite. It has a cell body located superficially (in layer 1) in the OE bearing apically located cilia and microvilli both, in a crypt-like invagination and longest axonal process extending towards the basal lamina.13,14,20,26,56 Their number in N. notopterus is less as compared to the other two types.
In the sensory region, two morphologically distinct types of OSNs, ciliated and microvillous are prevalent in teleosts.57 In addition to the teleosts, expression of both ciliated and microvillous sensory cells is seen in a primitive extant ray-finned fishes, the bichir (Polypterus senegalus and P. ornatipinnis58) and the sturgeons of genus Acipencer.59 They occur together but in varying proportions in different species.59 In the OE of N. notopterus, mOSNs visually seem to dominate over ciOSNs, same is observed in C. punctatus53 whereas ciOSNs are dominant over mOSNs in Labeo bata.60 In channel catfish, density of mOSNs is highest in the medial part of lamellae while that of ciOSNs is highest in the lateral areas of lamellae.61 In goldfish, mOSNs are most abundant in dorso-medial areas of the lamella close to the midline raphe.24 Third type of cells, crypt OSNs are also observed in N. notopterus but are very few in number. These are however widespread in number of teleost fishes.10,12,58,59,62-66 They are reported in Cyprinodonts,37 catfish, swordtail and needlefishes11 and in Zebrafish, Danio rerio.13
Expression of OSN polymorphism appears to follow an evolutionary pattern in those species that have been investigated. In an ancient jawless fish (superclass Agnatha), the sea lamprey (Petromyzon marinus), only ciliated OSNs are seen67,68 but three morphotypes are reported in P. marinus based on dendrite length and position of cell body within the OE.15 In elasmobranchs, these sensory cells bear only microvilli65,66,69-71 and crypt cell also appears in the OE.65,66 With the divergence of rayfinned fishes in different habitats OE seems to be populated by all the three OSNs.58,59,64 Overall, in fishes displaying all the three morphotypes, ciliated cells predominate with microvillous OSNs being fewer in number.16 Generally, crypt cells tend to be rare,63 and in some species, these do not appear in all the specimens10 and their number may also vary with season72 and sex.63 Ciliated OSNs are tuned toward bile salts and microvillous OSNs toward amino acids.16 However, recent electrophysiological studies concluded that ciliated OSNs might be termed as generalists which respond to varying species of odorants including amino acids, bile salts and other odorants whereas microvillous OSNs might be called as specialist, which respond specifically to amino acids and nucleotides.27,73
In addition to Agnatha15 and Elasmobranchs, OSN polymorphism has been reported in many Teleostomi such as bichir- Polypterus senegalus and P. ornatipinnis58 and sturgeon, Acipencer.59 In the teleosts it is reported in order Cypriniformes- goldfish, Carassius auratus,29 Aplochelius lineatus and Xiphophorus helleri;30 zebrafish, D. rerio,13 Siluriformes- channel catfish, Ictalurus punctatus,29,31 Salmonid fishes,16,32 and among Perciformes in round goby, Neogobius melanostomus.10
Order Osteoglossiformes is a representative of an early evolutionary lineage of teleost fishes. N. notopterus belongs to this order. In this fish, polymorphous OSNs are noted similar to those reported among the members of other teleosts including orders-cypriniformes, siluriformes and salmoniformes which are highly evolved orders. This indicates that OSN polymorphism is a conserved trait throughout the evolution of teleosts. Present study will be helpful to study the expression of receptor subtypes on each receptor cell and to know the physiological role assigned to each of the receptor cell types. Possibly these polymorphs respond to different odors, as seen in other teleosts.
This article complies with International Committee of Medical Journal editor’s uniform requirements for manuscript.
Conflict of Interests: None: Source of funding: None
Received Date : 05 March 2014; Revised Date : 09 April 2014; Accepted Date : 29 April 2014
References
1. Farbman AI. The cellular basis of olfaction. Endeavour 1994; 18: 2–8.
2. Valentinčič T. Olfactory discrimination in fishes. In: Reutter K, Kapoor BG, Ed., Fish chemosenses, Enfield (NH): Science Publishers 2005; 63–85.
3. Rajakaruna RS, Brown JA, Kaukinen KH, et al. Major histocompatibility complex and kin discrimination in Atlantic salmon and brook trout. Mol Ecol. 2006; 15: 4569–4575.
4. Olsén KH, Liley NR. The significance of olfaction and social cues in milt availability, sexual hormone status, and spawning behavior of male rainbow trout (Oncorhynchus mykiss). Gen Comp Endocrinol. 1993; 89: 107–118.
5. Stabell OB. Olfactory control of homing behaviour in salmonids. In: Hara TJ Ed., Fish chemoreception, London: Chapman and Hall 1992; 249–270.
6. Ueda H, Kaeriyama M, Mukasa K, et al. Lacustrine sockeye salmon return straight to their natal area from open water using both visual and olfactory cues. Chem Senses 1998; 23: 207–212.
7. Shoji T, Ueda H, Ohgami T, et al. Amino acids dissolved in stream water as possible home stream odorants for masu salmon. Chem Senses 2000; 25: 533–540.
8. Stacey NE, Sorensen PW. Reproductive pheromones. In: Sloman K, Balshine S, Wilson R, Ed., Behaviour and Physiology of Fish: Volume 24 (Fish Physiology), Academic Press 2006; 359–412.
9. Hansen A, Reutter K. Chemosensory systems in fish: structural, functional and ecological aspects. In: von der Emde G, Mogdans J, Kapoor BG, Ed., The Senses of Fish: Adaptations for the Reception of Natural Stimuli, Dordrecht Holland and Narosa Publishing House, New Delhi: Kluwer Academic Publishers 2004; 55–89.
10. Belanger RM, Smith CM, Corkum LD, et al. Morphology and histochemistry of the peripheral olfactory organ in the round goby, Neogobius melanostomus (Teleostei: Gobiidae). J Morphol. 2003; 257(1): 62–71.
11. Hansen A, Eller P, Finger TE, et al. The crypt cell: a microvillous ciliated olfactory receptor cell in teleost fishes. Chem Senses 1997; 22: 694–695.
12. Hansen A, Finger TE. Phyletic distribution of crypt-type olfactory receptor neurons in fishes. Brain Behav Evol. 2000; 55(2): 100–110.
13. Hansen A, Zeiske E. The peripheral olfactory organ of the zebrafish, Danio rerio: an ultrastructural study. Chem Senses 1998; 23(1): 39–48.
14. Hansen A, Zielinski BS. Diversity in the olfactory epithelium of bony fishes: development, lamellar arrangement, sensory neuron cell types and transduction components. J Neurocytol. 2005; 34(3–5): 183–208.
15. Laframboise AJ, Ren X, Chang S, et al. Olfactory sensory neurons in the sea lamprey display polymorphisms. Neurosci Lett. 2007; 414: 277–281.
16. Thommesen G. Morphology, distribution, and specificity of olfactory receptor cells in salmonid fishes. Acta Physiol Scand. 1983; 117(2): 241–249.
17. Yamamoto M, Ueda K. Comparative morphology of fish olfactory epithelium X. Perciformes, Beryciformes, Scorpaeniformes and Pleuronectiformes. J Fac Sci Tokyo Univ Sec IV Zool. 1979; 14: 273–297.
18. Hamdani EH, Alexander G, Døving KB. Projections of sensory neurons with microvilli to the lateral olfactory tract indicates their participation in feeding behavior in crucian carp. Chem Senses 2001a; 26: 1139–1144.
19. Morita Y, Finger TE. Differential projections of ciliated and microvillous olfactory receptor cells in the catfish, Ictalurus punctatus. J Comp Neurol. 1998; 398: 539–550.
20. Hamdani EH, Døving KB. The functional organization of the fish olfactory system. Prog Neurobiol. 2007; 82: 80–86.
21. Hamdani EH, Døving KB. Specific projection of the sensory crypt cells in the olfactory system in crucian carp, Carassius carassius. Chem Senses 2006; 31: 63–67.
22. Saito S, Yamamoto Y, Mori M, et al. Variety in histochemical characteristics of the olfactory receptor cells in a flatfish, barfin flounder (Verasper moseri). J Vet Med Sci. 2004; 66: 409–412.
23. Sato Y, Miyasaka N, Yoshihara Y. Mutually exclusive glomerular innervation by two distinct types of olfactory sensory neurons revealed in transgenic zebrafish. J Neurosci. 2005; 25(20): 4889–4897.
24. Hansen A, Anderson K, Finger TE. Differential distribution of olfactory receptor neurons in goldfish: structural and molecular correlates. J Comp Neurol. 2004; 477: 347–359.
25. Hamdani EH, Kasumyan A, Døving KB. Is feeding behaviour in crucian carp mediated by the lateral olfactory tract? Chem Senses 2001b; 26: 1133–1138.
26. Hamdani EH, Døving KB. The alarm reaction in crucian carp is mediated by olfactory neurons with long dendrites. Chem Senses 2002; 27: 395–398.
27. Hansen A, Rolen SH, Anderson K, et al. Correlation between olfactory receptor cell type and function in the channel catfish. J Neurosci. 2003; 23: 9328–9339.
28. Schmachtenberg O. Histological and electrophysiological properties of crypt cells from the olfactory epithelium of the marine teleost Trachurus symmetricus. J Comp Neurol. 2006; 495: 113–121.
29. Muller JF, Marc RE. Three distinct morphological classes of receptors in fish olfactory organs. J Comp Neurol. 1984; 222(4): 482–495.
30. Zeiske E, Melinkat R. Utrastructural studies on the epithelia of the olfactory organ of cyprinodonts (Teleostei, Cyprinodontoidea). Cell Tissue Res. 1976; 172: 245–267.
31. Caprio J, Raderman-Little R. Scanning electron microscopy of the channel catfish olfactory lamellae. Tissue Cell 1978; 10(1): 1–9.
32. Thommesen G. Specificity and distribution of receptor cells in the olfactory mucosa of char (Salmo alpines L.). Acta Physiol Scand. 1982; 115: 47–56.
33. Kluver H, Barrera E. A method for the combined staining of cells and fibers in the neurons system. J Neuropathalo Exp Neurol. 1953; 12: 400–403.
34. Nelson JS. Fishes of the World, 3rd ed. New York: John Wiley and Sons 1994; 1–600.
35. Schulte E, Riehl R. Feinstruktur der Regio olfactoria vom Piranha, Serrasalmus nattereri (Kner, 1860) (Teleostei, Characidae). Zool Anz Jena. 1978; 200: 119–131.
36. Schulte E. Untersuchungen an der Regio olfactoria des Aals, Anguilla Anguilla. L. Z Zellforsch. 1972; 125: 210–228.
37. Zeiske E, Kux J, Melinkat R. Development of the olfactory organ of oviparous and viviparous cyprinodonts (Teleostei). Z Zool Syst Evol. 1976; 14: 34–40.
38. Burne RH. The anatomy of the olfactory organ of teleostean fishes. Proc Zool Soc Lond. 1909; 2: 610–663.
39. Døving KB, Dubois-Dauphin M, Holley A, et al. Functional anatomy of the olfactory organ of fish and the ciliary mechanism of water transport. Acta Zoologica 1977; 58: 245–255.
40. Hara TJ. Olfaction in fish. Prod Neurobiol. 1975; 5: 271–335.
41. Kleerekoper H. Olfaction in Fishes. Bloomington: Indiana University Press 1969; 1–222.
42. Yamamoto M. Comparative morphology of the peripheral organ in teleosts. In: Hara TJ, Ed., Chemoreception in fishes, Amsterdam (The Netherlands): Elsevier 1982; 39–59.
43. Zeiske E, Theisen B, Breucker H. Structure, development and evolutionary aspects of the peripheral olfactory system. In: Hara TJ Ed., Fish Chemoreception, London: Chapman and Hall 1992; 13–39.
44. Khan FA, Jain MR, Saha SG, et al. FMRFamide-like immunoreactivity in the olfactory system responds to morphine treatment in the teleost Clarias batrachus: involvement of opiate receptors. Gen Comp Endocrinol. 1998; 110: 79–87.
45. Khan FA, Saha SG, Sarkar S, et al. β-endorphin-like immunoreactivity in the forebrain and pituitary of teleosts Clarias batrachus (Linn.). Gen Comp Endocrinol. 1999; 113: 290–301.
46. Biju KC, Singru PS, Schreibman MP, et al. Reproductive phase-related expression of GnRH-like immunoreactivity in the olfactory receptor neurons, their projections to the olfactory bulb and in the nervus terminalis in the female Indian major carp Cirrhinus mrigala (Ham.). Gen Comp Endocrinol. 2003; 133: 358–367.
47. Singru PS, Sakharkar AJ, Subhedar NK. Neuronal nitric oxide synthase in the olfactory system of an adult teleost fish, Oreochromis mossambicus. Brain Res. 2003; 977: 157–168.
48. Chakrabarti P. Histological and histochemical studies on the olfactory rosette of Mugil parsia (Hamilton). Folia Morphologica 2005; 64(1): 41–46.
49. Bhute YV, Masram SC, Raut IN, et al. Cytoarchitectonic pattern of the olfactory system, Forebrain and Pituitary gland of the Indian Major Carp, Labeo rohita. Annals Neurosci. 2007; 14: 30–36.
50. Bhute YV, Baile VV. Organization of the olfactory system of the Indian major carp Labeo rohita (Ham.): A scanning and transmission electron microscopic study. J Evol Biochem Physiol. 2007; 43: 342–349.
51. Ghosh SK, Chakrabarti P. Cellular architecture and functional aspects of the olfactory rosette of Wallago attu (Bleeker). Turk J Fish Aquat Sci. 2009; 9: 187–190.
52. Chakrabarti P, Guin S. Surface architecture and histoarchitecture of the olfactory rosette of freshwater lesser spiny eel, Macrognathus aculeatus (Bloch). Arch Pol Fish 2011; 19: 297–303.
53. Mandal DK, Roy D, Ghosh L. Structural organization of the olfactory epithelium of a spotted snakehead fish, Channa punctatus. Acta Ichthyol Piscat. 2005; 35(1): 45–50.
54. Patle PJ. Immunocytochemical studies on the effects of pesticide on olfactory-brain-pituitary-gonadal axis of a dwarf snakehead, Channa gachua (Hamilton). 2013; Ph.D. Thesis, RTM Nagpur University, Nagpur, M.S., India.
55. Baby SM, Ueck M, Prasada Rao PD. Gonadotropin–releasing hormone immunoreactive neurons and associated nicotinamide adenine nucleotide phosphate diaphorase–positive neurons in the brain of teleosts, Rhodeus amarus. Gen Comp Endocrinol. 2000; 120: 44–54.
56. Zielinski B, Hara TJ. Olfaction. In: Hara TJ, Zielinski BS, Ed., Sensory Systems Neuroscience: Volume 25 (Fish Physiology), Academic Press 2007; 1–43.
57. Zielinski B, Hara TJ. Morphological and physiological development of the olfactory receptor cells in rainbow trout (Salmo gairdneri) embryos. J Comp Neurol. 1988; 271(12): 300–311.
58. Zeiske E, Bartsch P, Hansen A. Early ontogeny of the olfactory organ in a basal actinopterygian fish: Polypterus. Brain Behav Evol. 2009; 73(4): 259–272.
59. Zeiske E, Kasumyan A, Bartsch P, et al. Early development of the olfactory organ in sturgeons of the genus Acipenser: a comparative and electron microscopic study. Anat Embryol. 2003; 206(5): 357–372.
60. Ghosh SK, Chakrabarti P. Distribution and organization of different cells lining the olfactory epithelium of the Indian minor carp, Labeo bata (Hamilton 1822): a light and scanning electron microscopic analysis. Pak J Biol Sci. 2011; 14: 736–741.
61. Erickson JR, Caprio J. The spatial distribution of ciliated and microvillous olfactory receptor neurons in the channel catfish is not matched by a differential specificity to amino acids and bile salt stimuli. Chem Senses 1984; 9: 127–141.
62. Castro A, Becerra M, Anadón R, et al. Distribution of calretinin during development of the olfactory system in the brown trout, Salmo trutta fario: comparison with other immunohistochemical markers. J Chem Neuroanat. 2008; 35(4): 306–316.
63. Bettini S, Lazzari M, Ciani F, et al. Immunohistochemical and histochemical characteristics of the olfactory system of the guppy, Poecilia reticulata (Teleostei, Poecilidae). Anat Rec. 2009; 292(10): 1569–1576.
64. Camacho S, Ostos-Garrido MV, Domezain A, et al. Study of the olfactory epithelium in the developing sturgeon. Characterization of the crypt cells. Chem Senses 2010; 35(2): 147–156.
65. Ferrando S, Bottero S, Pedemonte F, et al. Appearance of crypt neurons in the olfactory epithelium of the skate Raja clavata during development. Anat Rec. 2007; 290(10): 1268–1272.
66. Ferrando S, Gambardella C, Ravera S, et al. Immunolocalization of G-protein alpha subunits in the olfactory system of the cartilaginous fish Scyliorhinus canicula. Anat Rec. 2009; 292(11): 1771–1779.
67. Thornhill RA. The ultrastructure of the olfactory epithelium of the lamprey Lampetra fluviatilis. J Cell Sci. 1967; 2(4): 591–602.
68. VanDenbossche J, Seelye JG, Zielinski BS. The morphology of the olfactory epithelium in larval, juvenile and upstream migrant stages of the sea lamprey, Petromyzon marinus. Brain Behav Evol. 1995; 45(1): 19–24.
69. Takami S, Luer CA, Graziadei PP. Microscopic structure of the olfactory organ of the clearnose skate, Raja eglanteria. Anat Embryol. 1994; 190(3): 211–230.
70. Schluessel V, Bennett MB, Bleckmann H, et al. Morphometric and ultrastructural comparison of the olfactory system in ellasmobranchs: the significance of structure-function relationships based on phylogeny and ecology. J Morphol. 2008; 269(11): 1365–1386.
71. Ferrando S, Gallus L, Gambardella C, et al. G protein alpha subunits in the olfactory epithelium of the holocephalan fish Chimera monsterosa. Neurosci Lett. 2010; 472(1): 65–67.
72. Hamdani EH, Lastein S, Gregersen F, et al. Seasonal variations in olfactory sensory neurons – fish sensitivity to sex pheromones explained? Chem Senses 2008; 33(2): 119–123.
73. Sato K, Suzuki N. Whole-cell response characteristics of ciliated and microvillous olfactory receptor neurons to amino acids, pheromone candidates and urine in rainbow trout. Chem Senses 2001; 26: 1145–1156.