Annals of Neurosciences, Vol 12, No 2 (2005)
Annals of Neurosciences, Volume 12, Issue 2 (April), 2005
EXPRESSION OF PERCHLORIC ACID SOLUBLE 14KDA PROTEIN IN MOUSE BRAIN
#Corresponding author Biochemistry and Molecular
Biology Lab Department of Zoology Banaras Hindu University
Varanasi 221005 India
E-mail:
Abstract
Perchloric acid soluble protein (PSP), a member of the group of non-histone chromatin protein, consists of two identical subunits with a molecular mass of 14 kDa. It is highly conserved throughout the animal kingdom from prokaryotes to eukaryotes, reflecting its involvement in basic cellular regulation. It acts as an inhibitor for elongation step of protein synthesis. PSP expression has been reported to be tissue-specific, although it is predominantly present in liver and kidney cells. In this study, we report the expression of PSP in mice brain and compare it with liver. The expression of PSP was found to be much lower in brain than in liver, suggesting a tissue-dependent quantitative variation. The level of PSP mRNA was one-third and that of protein one-tenth in the brain as compared to the liver. Such discrepancy suggests that the brain PSP mRNA is less stable and its small fraction is translated leading to higher synthesis of other proteins in the brain.
Key Words: PSP, gene expression, mRNA
Introduction
Perchloric acid soluble protein (PSP) and PSP-like proteins are non-histone protein components of the chromatin, they can be extracted with 5% perchloric acid and 25% trichloroacetic acid. They were identified and co-extracted with histone HI and high mobility group (HMG) proteins from post-mitochondrial supernatant fraction of the rat liver (14). Initially the presence of PSP was reported only in the rat liver. Later PSP homologues were isolated from the goat liver (4,6), human monocytes (17), chicken liver (13), rat brain (.19) and pig liver (8). Baudier et al. (3) purified two perchloric acid soluble proteins, p43 and pl7, from the bovine brain. The p43 was identified as the neurospecific calmodulin-binding protein, neuromodulin.
PSP gene codes for 900bp mRNA having 411bp open reading frame (ORF), producing a protein of 137 amino acids. Nucleotide sequences of mammalian PSP cDNA show a high similarity to members of a new hypothetical family (YER057C/YJGF family) of small proteins (18). Structurally PSP is a homodimer consisting of two identical subunits with a molecular mass of $sl14 kda. Both polypeptide subunits are linked through disulfide bonds. Amino acid analysis revealed that the n-terminal amino acid sequence of PSP is acetylated (1,16).
Schmiedeknecht et al. (17) reported that the expression of 14.5kda PSP-like protein is significantly up-regulated upon cellular differentiation of human monocytes, in the rat kidney NRK-52E cells, the expression of PSP mRNA and protein is lower during the proliferating phase than the stationary phase. The expression pattern in proliferating cells is negatively correlated with the pattern of protein synthesis, i.e., when PSP expression is low, protein synthesis activity is high, and vice versa (9,10). The expression of PSP in rat hepatoma cell line DRLH84 was found to be less than 10% in tumorous part than in normal parts, suggesting that the expression of PSP is high in differentiated cells but low or down-regulated in the undifferentiated or dedifferentiated cells such as tumor cells (7). Thus it is evident that the expression of PSP is tissue-specific and varies at different cell stages during early development. In the present paper, comparison has been made for the expression of PSP between adult mice brain and liver. The expression of PSP mRNA and protein was found to be lower in brain than in liver, indicating tissue- dependent variation in PSP expression at adult age. Such variation suggests that the brain PSP mRNA is less stable and its only a small fraction translated leading to higher synthesis of other proteins in the brain.
Materials and Methods
Animal
AKR mice were maintained in a colony at 24 ± 2°c with free access to food and drinking water. They were exposed to 12h/ 12h light and dark schedule. Their average life span was 70 ± 5 weeks. Adult mice (25 weeks) were taken and sacrificed by cervical dislocation. The liver and brain of the mice were removed and used for measuring the expression of RNA and protein.
Preparation of tissue lysate
Liver and brain tissues of mice were homogenized separately in two volumes of lysis buffer consisting of cold 0.25M sucrose 50mM Tris.HCl pH 7.5, 25ml KCl and 10ml MgC12. The homogenate was centrifuged at 10,000Xg for 30 minutes. The supernatant was collected and the amount of protein in sample was measured according to Bradford (2) using BSA as a standard solution.
Western blotting
The protein in homogenate samples from liver and brain were analyzed on 15% Polyacrylamide gels according to the method of Laemmli (2). Following electrophoresis, the gel was transferred on to polyvinyl difluoride (PVDF) membrane (Sigma, USA) by semi-dry method using Hoefer transfer apparatus (Pharmacia Biosciences, USA). The blot was then incubated with PSP polyclonal antibody (1:2000 dilutions) for 2 hours on an orbital shaker at room temperature. Then it was washed in PBS three times for 5 min each. After washing, the membrane was incubated in HRPO-conjugated anti- rabbit IgG (1:4000 dilutions, Bangalore Genie, India) for 2 hours. The blot was again washed in PBS three times for 5 min each. After final washing, the signals were detected by ECL method (Pharmacia Biosciences, USA).
Slot blot analysis of PSP mRNA
Total RNA was isolated from the liver and brain (200mg each) of adult mice by Chomczynski and Sacchi method (5), A260 / A280 for each sample was calculated and found to be 1.7. RNA samples (15μ and 45μ) were loaded on to positively charged nylon membrane (Roche Applied Sciences, Germany) after assembling manifold slot- blot apparatus (Schleicher and Schuell, Germany) according to manufacturer's instruction. Then the membrane was air-dried and baked at 120°Cfor 30 minutes.
Preparation of hybridization probe
Plasmid containing PSP cDNA was isolated by alkaline lysis method (2) and digested with Eco RI. The purified cDNA fragment was labeled using DIG -11-dUTP (Roche Applied Sciences, Germany) as per the instruction manual.
The nylon membrane containing slot blotted RNA was sealed in a plastic bag containing pre-hybridization solution (7% SDS, 50ml sodium phosphate buffer, pH 7.0,50% formamide, 2% blocking reagent, 50μl/ml denatured salmon sperm DNA, 5x SSC, 0.1% N laurylsarcosine) and allowed to pre-hybridize overnight at 50°C. The membrane was then taken out from the pre-hybridization solution and placed in hybridization solution containing 25ng labeled cDNA/ml in a sealed plastic bag and allowed to hybridize overnight at 50°C. After completion of hybridization, washing and detection was done according to the method mentioned in the DIG system User's Guide for Filter Hybridization (Roche Applied Sciences, Germany).
QUANTIFICATION OF DATA
Signals obtained from western blot and slot blot were densitometrically quantified by Flurochem software version 2 (Alpha Innotech. Corp, USA).
Results
Western blot analysis of PSP protein
Amount of total protein in mice liver and brain sample was measured by Bradford method (2). One unit of A595 for BSA was found equivalent to 112.5mg pf protein. Protein yield in the liver and brain sample was found to be 92.80mg and 29.70mg/ gm wet tissue weight, respectively. Equal amount of crude protein (50 mg) from brain and liver were electrophoresed on 15% SDS-polyacrylamide gel (Fig 1) and transferred onto PVDF membrane for western blot analysis. In the western blot, one =sl14 kDa band was seen. PSP from mice brain and liver showed strong cross-reaction with rat anti PSP antibody. Expression of PSP protein was found to be ∼10 times lower in brain when compared to liver (Fig.2).
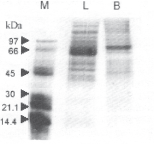
Fig. 1. Coomassie brilliant blue sds-page of liver and brain extract of adult mouse: 50 μg protein from liver (L) and brain (B) was loaded on 15% Polyacrylamide gel containing 0.1% SDS. After completion of electrophoresis, protein bands were visualized by staining with coomassie brilliant blue. (M, size marker represented in kDa)
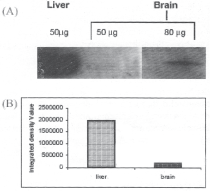
Fig. 2. Western blot analysis of psp protein: (a) western blot of psp protein from liver (50μg) and brain (50 μg and 80 μg), (b) densitometric graph of psp protein expression from liver (50 μg) and brain (50 μg).
Slot-blot hybridization analysis of PSP mRNA
Purified total RNA samples of mice liver and brain were resolved on 1% Agarose-formaldehyde gel. The integrity of RNA was found to be as represented by bands of 28S and 18S RNA (Fig.3). Thereafter RNA was slot-blotted onto nylon membrane and hybridized with DIG-11-dUTP labeled PSP cDNA. Densitometrie result (Fig.4) showed that the expression of PSP mRNA was three times lower in brain as compared to liver.

Fig. 3. 1% Agarose-formaldehyde gel electrophoresis of total RNA: 10μg RNA obtained from mice liver and brain were loaded to check integrity of RNA. 28s and 18s RNA are shown by arrows.
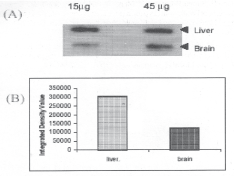
Fig. 4. Slot blot analysis of PSP mRNA: (A) Slot blot of mRNA from mice liver and brain. 15μg and 45μg RNA was loaded. Slot blot was probed with rat PSP cDNA labeled with Dig-11-dUTP (B) Histogram representing densitometric scanning of signals from 15jiig RNA slot of mice liver and brain.
Discussion
In the present study, the expression pattern of PSP m (RNA) and protein was compared between adult mice brain and liver. We used mice brain as a model organ and liver was used as an organ of reference for the expression of PSR In both brain and liver, a14kda band was obtained by immuno detection. For this study, antibody directed against rat liver PSP was used. The cross reactivity of this antibody with mice liver and brain PSP suggested that these proteins are highly conserved among different species. Oka et al. and others (7,8,15–17) confirmed that PSP is conserved from bacteria to higher organisms such as mice, rat, pig and human.
PSP inhibits protein synthesis (1) in a concentration-dependent manner. It acts as an inhibitor for the elongation stage of translation. The endoribonuclease activity of PSP causes disaggregation of polysomes through fragmentation of polysomal mRNA (14). The recombinant PSP suppresses the production of immunoglobulin in human hybridoma hb4c5 cells (11).
The results of our study showed that PSP mRNA and protein are less expressed in mouse brain than the liver. The level of brain PSP mRNA was one-third and protein one-tenth as compared to the liver. This variation may be attributed to either less copy number or short half-life of PSP mRNA. Furthermore, it appears that all mRNA molecules in the brain are not translated, suggesting the regulation of PSP expression at both transcriptional and translational steps.
The expression of PSP changes within the same tissue at different differentiation states of the cell and developmental stages of the animal. In the rat brain, PSP expression increased gradually from the first day to 60th day. Suzuki et al. (19) reported that the expression of brain PSP at 60th day was two times higher than. that at the first day. PSP was expressed in the entire brain regions including frontal cortex, posterior cortex, cerebellum, hippocampus, olfactory bulb, striatum, thalamus, mid brain, entorhinal cortex, pons, medulla and spinal cord. Immunohistdchemical study showed that the rat brain psp was expressed in ependymal cells of choroid plexus and glial cells of other regions.
In our present study, the expression of PSP was found to be much lower in brain than in liver, suggesting a tissue- dependent quantitative variation. Such discrepancy suggests that brain PSP mRNA is less stable and its only a small fraction is translated leading to higher synthesis of other proteins in the brain. This may have a great significance in the cellular physiology of brain. The inhibition of translation by PSP may help in tight regulation of protein level in cells.
References
1. Asagi K, Oka T, Arao K, Suzuki I, Thakur MK, Izumi K, Natori Y. Purification, characterization and differentiation dependent expression of a perchloric acid soluble protein from rat kidney. Nephron 1998; 79:80–90.
2. Ausubel FM, Brent R, Kinston RE, Moore DT, Siedman JG, Smith JA, Struthl K. Short protocols in molecular biology John Wiley & Sons, New York 1995.
3. Baudier J, Bronner C, Kligman D, Cole RD. Protein kinase C substrates from bovine brain: Purification and characterization of neuromodulin, a neuron-specific calmodulin-binding protein. J. Biol. Chem. 1989; 264: 1824–1828.
4. Ceciliani F; Faotto L, Negri A, Colombo, Berra B, Bartorelli A, Ronchi S. The primary structure of UK-114 tumor antigen. FEBS Lett. 1996; 893:147–150.
5. Chomczynski P, Sacchi N. Single step method of ma isolation by acid guanidium thiocyanate- phenol-chloroform extraction. Anal Biochem. 1987; 162: 156–159.
6. Ghezzo F, Berta GN, Bussolati B, Bosio A, Corvetti G, Di Carlo F, Bussolati G, Guglielmone R, Bartorelli A. Perchloric acid soluble proteins from goat liver inhibit chemical carcinogenesis of Syrian hamster check pouch carcinoma. Br. J. cancer 1999; 79: 54–58.
7. Kaneki K, Kanouchi H, Matsumoto M, Kawasaki Y, Akuzawa M, Oka T. Down regulation of a novel protein, psp in rat hepatoma cell drlh 84-beared tumor. J Vet. Med. Sci. 2003; 65: 781–785.
8. Kaneki K, Matsumoto M, Suzuki K, Akuzawa M, Oka T. Purification, characterization and developmental expression of pig liver psp. comp. Biochem. Physiol. B Biochem. Mol. Bio. 2003; 134: 571–578.
9. Kanouchi H, Oka T, Agasi K, Tachibana H, Yamada K. Expression and cellular distribution of perchloric acid soluble protein is dependent on the cell proliferating states of NRK- 52e cells. Cell. Mol. Life Sci. 2000; 57:1103–1108.
10. Kanouchi H, Oka T, Tachibana H, Yamada K. Recombinant expression of perchloric acid soluble protein reduces cell proliferation. Cell. Mol. Life Sci. 2001; 58:1340–1343.
11. Kanouchi H, Matsuo A, Oka T, Tachibana H, Yamada K. Recombinant perchloric acid-soluble protein suppresses the immunoglobulin production of human-human hybridoma hb4c5 cells, in vitro cell dev biol – animal 2003; 39: 263–265.
12. Morishita R, Kawagoshi A, Sawasaki T, Madin K, Ogasawara T, Oka T, Endo Y Ribonuclease activity of rat liver perchloric acid soluble protein, a potent inhibitor of protein synthesis. J. Bio. Chem. 1999; 274: 20688–20692.
13. Nordin H, Matsomoto M, Suzuki K, Kaneiki K, Natori Y, Kishi K, Oka T. Purification, expression and developmental expression of chick (gallus domesticus) liver psp protein, comp, biochem. Physiol. B Biochem. Mol. Biol. 2001; 128: 135–143.
14. Oka T, Tsuji H, Noda C, Sakai K, Hongs Y, Suzuki I, Munoz S, Natori Y Isolation and characterization of a novel perchloric acid soluble protein inhibiting cell free protein. J. Biol. Chem. 1995; 270: 30060–30067.
15. Oka T, Nishimoto Y, Sasagawa T, Kanouchi H, Kawasaki Y, Natori Y Production of functional rat liver psp protein in e. coli. Cell Mol. Life. Sci. 1999; 55: 131–134.
16. Sasagawa T, Oka T, Tokumura A, Nishimoto Y, Munoz S, Kuwahata M, Okita M, Tsuji H, Natori Y, Analysis of the fatty acid components in a perchloric acid soluble protein, biochim. Biophys. Acta 1999; 1437: 317–324.
17. Schmiedeknecht G, Kerkhoff C, Orso E, Stohr J, Aslanidis C, Nagy GM, Knuechel R, Schmitz G. Isolation and characterization of 14.5 kda trichloro acetic acid soluble translational inhibitor protein from human monocytes that is up regulated upon cellular differentiation. Eur. J. Biochem. 1996; 242: 339–351.
18. Sinha S, Rappu P, Lange SC, Mäntsälä FJ Zalkin H, Smith JL. Crystal structure of bacillus subtilis yabj, a purine regulatory protein and member of the highly conserved yjgf family. Proc. Natl. Acad. Sci. Usa 1999; 96: 13074–13079.
19. Suzuki K, Nishida T, Matsumoto M, Kanouchi H, Kaneiki K, Oka T. Purification, characterization and developmental expression of rat brain psp protein. Biochim. Biophys Acta 2001; 1527: 47–53.
(c) Annals of Neurosciences.All Rights Reserved