Annals of Neurosciences, Vol 13, No 2 (2006)
Annals of Neurosciences, Volume 13, Issue 2 (April), 2006
MELATONIN COUNTERS THE 5-FLUOROURACIL-INDUCED DECREASE IN BRAIN SEROTONIN AND DOPAMINE LEVELS
Corresponding author
Daya S
Faculty of Pharmacy, Rhodes University, Grahamstown 6140, South Africa,
Abstract
It has been demonstrated that both 5-fluorouracil and melatonin inhibit rat liver tryptophan-2,3-dioxygenase activity. This study sought to examine the effects that this inhibition has on the levels of various neurotransmitters in the brain. By inhibiting TDO, melatonin increases brain serotonin levels. Melatonin also increases dopamine levels but does not alter norepinephrine levels. 5-Fluorouracil has been found to decrease brain levels of all three neurotransmitters (i.e. serotonin, norepinephrine and dopamine). This could be due to the antineoplastic inhibiting DNA synthesis and gene expression of the enzymes required in the synthesis of these neurotransmitters. The decrease in norepinephrine levels induced by 5-fluorouracil could also be due to decreases in serotonin and dopamine levels. Dopamine can be converted to norepinephrine, whose release can be induced by serotonin. Even though 5-fluorouracil and melatonin alter brain serotonin levels, neither drug alters the conversion of serotonin to the metabolite 5-hydroxy-indole acetic acid. The same is true for the dopamine metabolite 3,4-dihydroxyphenylacetic acid. By decreasing the levels of norepinephrine, serotonin and dopamine, 5-fluorouracil could contribute to the development or exacerbation of depression in cancer patients. Melatonin, if administered together with 5-fluorouracil, could counter this 5-fluorouracil-induced depression by significantly increasing brain serotonin and dopamine levels. The use of melatonin as adjunctive therapy with 5-fluorouracil to improve patients' quality of life needs to be investigated further.
Key words : Tryptophan-2,3-dioxygenase activity, Neurotransmitter levels, Depression
Introduction
The enzyme tryptophan-2,3-dioxygenase (TDO) exists in both a holoenzyme and apoenzyme form (1). An increase in the activity of TDO in the liver results in a decrease in the availability of circulating tryptophan that can enter the brain (2). Once in the brain, tryptophan is converted to the neurotransmitter serotonin (5-HT) (2–4). Daya et al (5) thus conclude that there is an inverse relationship between the levels of 5-HT in the brain and hepatic TDO. TDO activity is a key peripheral indicator of the cerebral availability of tryptophan (6). 5-HT, in turn, has been found to have an effect on the levels of other neurotransmitters in the brain, such as norepinephrine (NE) (7).
According to the biogenic amine hypothesis of depression, this disorder is caused by a deficiency of the monoamine neurotransmitters 5-HT and NE (8–10). The dopamine hypothesis suggests that low levels of dopamine (DA) also contribute to this disorder (11,12). The kynurenine hypothesis states that increased TDO activity leads to increased levels of some of the metabolites of tryptophan, such as kynurenine, and that these may be responsible for depression (13, 14). Since melatonin has antidepressant properties (15), these authors (16) have recommended that the hormone be used to improve the quality of life of cancer patients who are on 5-FU therapy.
Materials and Method
Male Wistar rats weighing between 200g and 300g were used. 5-fluorouracil, melatonin and haematin were purchased from Sigma Chemical Company, St Louis, MO, USA. All the other chemicals used were obtained from local suppliers and were of the highest purity available. All solutions were prepared using de ionized water (Milli R/Q System, Millipore® ), and melatonin was dissolved using absolute ethanol as a co solvent in a final concentration of 1%. Four groups, each group containing 5 rats, were used. The animals were subjected to a diurnal lighting system of 12 hours of light alternating with 12 hours of darkness. The animals had access to water and food ad libitum. The injection volumes of saline, 5-FU and melatonin were 0.5 ml, while the injection volume of haematin, dissolved in saline, was 0.1 ml. All injections were given intraperitoneally. A 5-day treatment protocol was used. The control group received 0.9% NaCl at 08h00, 12h00 and 16h00. The 5-FU group received 1 mg/ kg 5-FU and the melatonin group received 1 mg/kg melatonin at the same times. The 3rd group of rats received both 1 mg/kg 5-FU and 1 mg/kg melatonin at the same times. All groups received haematin at a dose of 1 mg/kg at 09h30 and 17h30. On the sixth day, the rats were killed by cervical dislocation and the livers were removed immediately, perfused with 0.9% NaCl and assayed for TDO activity (17). The forebrains were dissected at the time of sacrifice and stored in liquid nitrogen at -70°C until the neurotransmitter levels were determined. This was done as described by Maharaj et al (3); the levels of NE, 5-HT, dopamine (DA), 3,4-dihydroxyphenylacetic acid (DOPAC) and 5-hydroxyindole acetic acid (5-HIAA) were determined. The analysis was performed on a high performance liquid chromatographic (HPLC) system. The potential was +0.74 V; the range was 5 nA and the total baseline was +0.43 nA. The oxidation potential was positive. The flow rate was 0.7 ml/ min and the mobile phase used was a combination of: 8.65 M heptane sulphonic acid, 0.27 mMEDTA, 13% acetonitrile, 0.43% triethylamine and 0.22% o-phosphoric acid. The column used was a C18 one, with the dimensions 4.6 × 250 mm, the injector was a Rheodyne® (USA) one and the column pressure was 1498 psi.
Results
Both 5-fluorouracil and melatonin were found to inhibit TDO activity. 5-FU inhibits only the apoenzyme, whereas melatonin inhibits both the apoenzyme and holoenzyme, as reflected in Figure 1 As shown in table 1, 5-fluorouracil reduced NE levels significantly from 8.58 pmol/mg to 5.87 pmol/mg, while melatonin did not change this. When both drugs are administered together, NE levels are significantly higher compared to when 5-FU is administered alone, but lower than the control levels. Although both drugs alter 5-HT levels, neither drug alters 5-HIAA levels. While melatonin alone increases 5-HT levels from 0.43 pmol/mg to 0.54 pmol/mg, 5-FU alone causes a decrease to 0.28 pmol/mg. The administration of both drugs in combination results in 5-HT levels rising to levels greater than with melatonin alone. Both 5-FU and melatonin alone do not alter DOPAC levels. Melatonin induces a significant rise in DA levels from 1.13 pmol mg to 3.83 pmol/mg, while 5-FU reduced this to 0.59 pmol/mg. Those animals which received both drugs showed DA levels of 1.23 pmol/mg. Thus the concomitant administration of melatonin prevents the reduction by 5-FU and increases DA to levels greater than the control values.
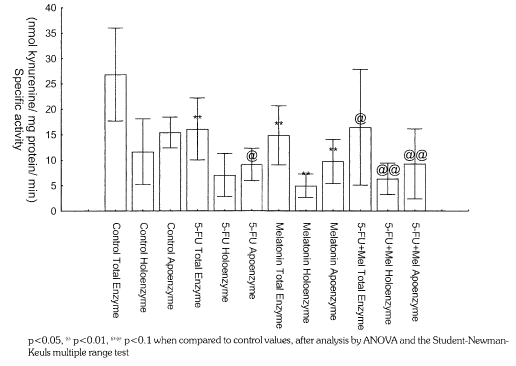
Figure I: The effect of 5-FU and melatonin on the specific activity of rat liver TDO in vivo
Discussion
An inverse relationship exists between cerebral 5-HT and hepatic TDO levels (5). Since both 5-FU and melatonin have been found to inhibit TDO, it is expected that the neurotransmitter analysis would reflect increased 5-HT levels. NE levels are expected to increase as well, since 5-HT is known to cause the release of this neurotransmitter (7). It was found that melatonin increases both 5-HT and NE levels. This is in accordance with the findings of Anton-Tay et al (18) and Wendel et al (19). These increased neurotransmitter levels could be due to inhibition of MAO by melatonin (20). 5-FU, unexpectedly, caused significant decreases in these neurotransmitter levels, whereas melatonin blunted the 5-FU-induced decrease in NE and 5-HT levels. 5-HIAA levels were not affected by either drug, suggesting that neither 5-FU nor melatonin alter the main route of the metabolism of 5-HT.
A possible explanation for 5-FU inhibition of TDO combined with a decrease in 5-HT levels is in considering 5-FU's action as an antimetabolite disrupting DNA synthesis. The biosynthesis of 5-HT requires enzymes and this is ultimately controlled at the level of gene expression and protein synthesis. 5-FU is known to enter the brain and can thus disrupt 5-HT synthesis in the brain by affecting gene expression. The same could be true for NE and DA as well (21).
| Group | Concentration (pmol/mg tissue) n=5 | ||
|---|---|---|---|
| NE | 5-HIAA | 5-HT | |
| Control | 8.58 ± 0.5 | 0.93 ± 0.064 | 0.43 ± 0.031 |
| Melatonin | 8.36 ± 0.12 | 0.97 ± 0.018 | 0.54 ± 0.028*,@ |
| 5-FU | 5.87 ± 0.16* | 0.93 ± 0.032 | 0.28 ± 0.023*,@ |
| Melatonin + 5-FU | 7.72 ± 0.47* | 0.94 ± 0.078 | 0.57 ± 0.062* |
* p<0.05 compared to control; @ p<0.01 compared to Melatonin + 5-FU. ANOVA, followed by the Student-Newman-Keuls multiple range test, was used for statistical analysis.
| Group | Concentration (pmol/mg tissue) n=5 | |
|---|---|---|
| DOPAC | DA | |
| Control | 0.94 ± 0.05 | 1.13 ±0.074 |
| Melatonin | 0.86 ± 0.047 | 3.83 ± 0.21@@ |
| 5-FU | 0.52 ± 0.047 | 0.59 ± 0.12@,** |
| Melatonin + 5-FU | 0.84 ± 0.21 | 1.23 ± 0.036 |
@ p<0.01, @@ p<0.001 compared to control; ** p<0.001 compared to Melatonin + 5-FU. ANOVA, followed by the Student-Newman-Keuls multiple range test, was used for statistical analysis.
DA levels increased by the administration of melatonin and were reduced by 5-FU. The enzyme dopamine β-hydroxylase catalyses the formation of NE from the catecholamine DA (22); increased levels of DA would thus be expected to be associated with increased levels of NE. Melatonin, which significantly increased DA levels, had no significant effect on NE levels. 5-FU, though, markedly reduced NE levels. One mechanism for this may be the decrease in DA levels due to 5-FU. 5-FU also decreases 5-HT levels, thereby further reducing NE levels. Neither drug alters DOPAC levels; DOPAC is a metabolite of DA and this finding indicates that neither 5-FU nor melatonin affect the metabolism of DA (23).
Lissoni has suggested that melatonin to be used as adjunctive therapy with 5-FU, to enhance the efficacy and reduce the toxicity of the latter to improve patients' quality of life (16). The findings of this study support this, in that melatonin was found to increase the levels of 5-HT and DA. When administered with 5-FU in this study, 5-HT and DA levels were increased to levels greater than the control. Melatonin has been found to have antidepressant properties (15), and its ability to increase the levels of these neurotransmitters may be one of the mechanisms by which it achieves this.
Acknowledgement
LC is a holder of an Andrew Mellon Foundation Scholarship and a National Research Foundation Scholarship.
References
1. Feigelson P, Greengard O. A microsomal iron-porphyrin activator of rat liver tryptophan pyrrolase. J Biol Chem 1961; 236: 153–157.
2. Maharaj H, Maharaj DS, Saravanan KS, Mohanakumar KP Daya S. Aspirin curtails the acetaminophen-induced rise in brain norepinephrine levels. Metabolic Brain Disease 2004; 19(1/2): 81–87.
3. Fernstrom JD, Wurtman RJ. Brain serotonin content: physiological dependence on plasma tryptophan levels. Science 1971; 173: 149–152.
4. Carlsson A, Lindqvist M. Dependence of 5-HT and catecholamine synthesis on concentrations of precursor amino-acids in rat brain. Naunyn Schmiedebergs Arch Pharmakol 1978; 303: 157–164.
5. Daya S, Nonaka KO, Buzzel GR, Reiter RJ. Heme precursor 5-aminolevulinic acid alters brain tryptophan and serotonin levels without changing pineal serotonin and melatonin concentrations. J Neurosci Res 1989; 23: 304–309.
6. Badawy AA-B, Welch N, Morgan CJ. Tryptophan pyrrolase in heme regulation: the mechanism of the opposite effects of tryptophan on rat liver 5-aminolaevulinate synthase activity and the haem saturation of tryptophan pyrrolase. Biochemical Journal 1981; 198: 309–314.
7. Xi-Ming Li S, Perry KW, Wong DT. Influence of fluoxetine on the ability of bupropion to modulate extracellular dopamine and norepinephrine concentrations in three mesocorticolimbic areas of rats. Neuropharmacology 2002; 42: 181–190.
8. Lapin IP, Oxenkrug GF. Intensification of the central serotonergc processes as a possible determinant of the thymoleptic effect. Lancet 1969; 1: 132–136.
9. Bunney WE Jr, Davis JM. Norepinephrine in depressive reactions: a review. Arch Gen Psychiat 1965; 13: 483.
10. Schildkraut JJ. The catecholamine hypothesis of affective disorders: a review of supporting evidence. A J Psych 1965; 122: 509.
11. Blier P de Montigny C. Current advances and trends in the treatment of depression. TIPS 1994; 15: 220.
12. Syvälahti EKG. Biological aspects of depression. Acta Psychiatr Scand 1994; 89(377): 11.
13. Curzon G, Bridges PK. Tryptophan metabolism in depression. J Neurol Neurosurg Psychiatry 1970; 33: 698.
14. Curzon G. Tryptophan pyrrolase – a biochemical factor in depressive illness. British J Psychiatry 1969; 115: 1367–1374.
15. Lissoni P, Barni S, Meregalli S et al. Modulation of cancer endocrine therapy by melatonin: a phase II study of tamoxifen plus melatonin in metastatic breast cancer patients progressing under tamoxifen alone. British J Cancer 1995; 71: 854–856.
16. Lissoni P, Barni S, Mandala M, Ardizzoia A, Paolorossi F, Vaghi M, Longarini R, Malugani F Tancini G. Decreased toxicity and increased efficacy of cancer chemotherapy using the pineal hormone melatonin in metastatic solid tumour patients with poor clinical status. European J Cancer 1999; 35: 1688–1692.
17. Badawy AA-B, Evans M. Regulation of rat liver tryptophan pyrrolase by its cofactor haem: Experiments with haematin and 5-aminolaevulinate and comparison with the substrate and hormonal mechanisms. Biochemical Journal 1975; 150: 511–520.
18. Anton-Tay F, Chou C, Anton S, Wurtman RJ. Brain serotonin concentration: elevation following intraperitoneal administration of melatonin. Science 1968; 162(3850): 277–278.
19. Wendel OT, Waterbury LD, Pearce JA. Increase in monoamine concentration in rat brain following melatonin administration. Experientia 1974; 30: 1167–1170.
20. Urry RL, Ellis LC. Monoamine oxidase activity of the hypothalamus and pituitary: alterations after pinealectomy, changes in photoperiod, or additions of melatonin in vitro. Experientia 1975; 31(8): 891–892.
21. Dollery C. 5-Fluorouracil. Ed: Therapeutic drugs. United Kingdom. Churchill Livingstone, Longman Group UK, 1991; 1: F68–73.
22. Menniti FS, Diliberto EJ Jr. Newly synthesised dopamine as the precursor for norepinephrine synthesis in bovine adrenomedullary chromaffin cells. J Neurochemistry 1989; 53(3): 890–897.
23. Zetterstrom T, Sharp T, Collin AK, Ungerstedt U. In vivo measurement of extracellular dopamine and DOPAC in rat striatum after various dopamine-releasing drugs; implications for the origin of extracellular DOPAC. European J Pharmacol 1988; 148(3): 327–334.
(c) Annals of Neurosciences.All Rights Reserved