Annals of Neurosciences, Vol 13, No 2 (2006)
Annals of Neurosciences, Volume 13, Issue 2 (April), 2006
REVERSAL OF DIAZEPAM TOLERANCE AND WITHDRAWAL-INDUCED HYPER LOCOMOTOR ACTIVITY AND ANXIETY BY MELATONIN IN MICE
Corresponding author
Dr. S.K. Kulkarni
Professor of Pharmacology, University Institute of
Pharmaceutical Sciences, Panjab University,
Chandigarh -160 014, India
(+91–172–534113, 541142, 779426 Fax: +91–172–779426E-
Mail:
Abstract
Background & Objectives: Earlier studies from our laboratory have confirmed the role of melatonin in the reversal of morphine tolerance and dependence in mice. The present study was performed to explore the possible involvement of melatonin in the reversal of diazepam tolerance and dependence in mice.
Methods: Diazepam (20 mg/kg/day, i.p.) was administered chronically on days 1–21. Mirrored-chamber was used to evaluate the anxiogenic reaction in mice due to withdrawal. Melatonin (2.5 or 5mg/kg; i.p.) was administered daily prior to diazepam administration for 21 days.
Results: Chronic administration of diazepam (20 mg/kg/day, i.p.) on days 1–21 and its withdrawal produced anxiogenic reaction in mice as assessed in the mirrored-chamber test. Daily administration of melatonin (2.5 or 5 mg/kg, i.p.) prior to diazepam for 21 days prevented withdrawal-induced anxiety in mice. However, acute administration of a single dose of melatonin (2.5 or 5 mg/kg), to animals withdrawn from diazepam, i.e. on the 22nd day, did not prevent withdrawal-induced anxiety. Diazepam withdrawal also induced a significant increase in the locomotor activity of mice indicating an anxiogenic response. Daily administration of melatonin (2.5 or 5 mg/kg) prior to diazepam for 21 days also prevented withdrawal- induced increased locomotor activity. Both acute and chronic administration of melatonin (2.5 and 5 mg/kg) exhibited a significant protection against diazepam withdrawal-induced anxiety and hyper locomotor activity in mice.
Conclusions: The result suggests the protective effect of this safe drug, melatonin, in the management of diazepam withdrawal reactions.
Key words: Diazepam dependence, Diazepam tolerance, Melatonin, Withdrawal jumps
Introduction
The benzodiazepines are widely prescribed for the treatment of anxiety and sleep disorders. These agents are extremely safe, but tolerance (1–3) and physical dependence develops rapidly upon long-term exposure (1–5). In animal models tolerance to their anxiolytic activity can also be measured. On abrupt withdrawal from benzodiazepine exposure, patients can experience a number of symptoms indicative of a dependent state and physical aspects of the withdrawal phenomena can be reproduced in animals. The benzodiazepines produce their beneficial effects by interaction with GABA A receptors, but it appears that tolerance and dependence are not due to a simple down regulation of these receptors (6).
Pineal hormone melatonin has been suggested to produce its CNS depressant effects by interacting with the benzodiazepine receptors (7). Further, the presence of high affinity melatonin recognition sites in the CNS, particularly their localization in areas such as forebrain and spinal cord, show that these receptors may be involved in the regulation of sensory transmission, visceral and autonomic reflexes (8,9). Melatonin (N-acetyl-5-methoxy-tryptamine), the chief indolamine produced by the pineal gland, has been shown to be an effective antioxidant and free radical scavenger (10–13). Melatonin is a broad spectrum free radical scavenger and indirect antioxidant. Melatonin, apart from direct free radical scavenging activity, indirectly enhances antioxidative defense mechanisms by increasing the activities of several antioxidant enzymes and by stimulating the synthesis of another important intracellular antioxidant, glutathione (GSH) may be involved in the regulation of sensory transmission, visceral and autonomic reflexes (14–15). Melatonin, because of its small size and high lipophilicity, crosses biological membranes easily, thus reaching all components of the cell (16).
Melatonin has been shown to exert an antioxidant effect on dopaminergic neurons (17), as well as dopaminergic modulating activities (18) suggesting thereby the possibility of melatonin being useful in the treatment of some aspects of drug abuse in man. In the present study, therefore, a possible effect of melatonin in the induction and expression of benzodiazepine tolerance and withdrawal-induced hyperlocomotor activity and anxiety was studied.
Materials and Methods
Animals
Albino mice (Laka strain) of either sex (20–25 g, bred in Central Animal House facility of the Panjab University, Chandigarh) were housed 5 per cage at room temperature and allowed to adapt to laboratory conditions for at least 2 days before the initiation of any experiment. The animals were housed under a natural light and dark cycle, and had free access to food and water. Each animal was used only once. All experiments were carried out between 0900 and 1700 h. The experimental protocols were approved by the Institutional Animal Ethics Committee and conducted according to the Indian National Science Academy Guidelines for the use and care of experimental animals.
Induction of BZD tolerance and withdrawal
Animals were made BZD dependent by administration of diazepam (20 mg/kg/day, i.p.) for 21 consecutive days (19,20). The injections were made daily at 1000 h. Control animals were injected with the vehicle alone. Development of BZD tolerance was assessed 30 min after the last injection of diazepam, and the spontaneous drug withdrawal was started 24h after the last injection. Similarly, in combination studies, melatonin and diazepam were administered concurrently for 21 days. The control animals were treated with the vehicle alone.
Treatment schedule
Diazepam (20 mg/kg, i.p.) was administered once daily for 21 days. Melatonin (2.5 or 5 mg/kg, i.p.) was administered for 21 days. Similarly, in combination studies melatonin and diazepam were administered concurrently for 21 days. For the assessment of effect of melatonin on the expression of tolerance and withdrawal, mice that had received diazepam during the induction phase (days 1–21) were acutely treated with melatonin on day 22. The control animals were treated with vehicle alone. Various treatment groups (pretreatment : treatment) included (1) saline : vehicle; (2) vehicle : diazepam (20 mg/kg, i.p.); (3) saline : melatonin (2.5 or 5 mg/kg, i.p.); (4) melatonin (2.5 or 5 mg/kg, i.p.) : diazepam (20 mg/kg, i.p.).
Measurement of locomotor activity
The diazepam withdrawal-induced hyper loco motor activity of mice was measured in actophotometer for a period of 5 minutes and the ambulatory and total activities recorded. The locomotion was expressed in terms of total photobeam counts/5 min. per animal.
Measurement of anxiety – Mirrored chamber test
The mirrored chamber used for mice consists of a wooden chamber having a mirror-chamber enclosed within it. During the 5 min test session the following parameters were noted: (i) transfer latency, (ii) the total time spent in mirror-chamber, (iii) number of entries the animal made in mirror-chamber. Animals were put individually at the distal corner of the mirrored chamber facing towards the mirror-chamber at the beginning of the test. An anxiogenic response was defined as decreased number of entries and time spent in the mirror chamber (21).
Drugs
Diazepam (Intas, India) was dissolved in distilled water (water soluble injection). Melatonin (Morepen, India) was dissolved in distilled water. The selection of doses was based on previous work done in our pharmacology research laboratory.
Statistical Analysis
The data expressed as mean ± SEM were analyzed by one way analysis of variance (ANOVA) followed by Dunnett's t-test. A value of P < 0.05 was considered statistically significant.
Results
Effect of melatonin on diazepam tolerance and withdrawal in mice on chronic administration of diazepam (20 mg/kg/day, i.p.) resulted in tolerance to both the behavioral sedation and anxiolytic effects. Abrupt discontinuation of diazepam, after 21 days treatment, resulted in withdrawal syndrome manifested as hyper loco motor activity and severe anxiety, i.e., a decrease in the number of entries and time spent in mirrored chamber at 24, 48 and 72h after the last injection. The peak withdrawal hyper loco motor activity and anxiety were observed on the third day of diazepam withdrawal. Co-administration of melatonin (2.5 or 5 mg/kg, i.p.) significantly prevented the development of diazepam tolerance and attenuated the withdrawal-induced hyper locomotion and anxiety state in mice. No significant alterations in any parameter were observed with melatonin (2.5 or 5 mg/kg, i.p.) given alone for 21 days when compared with vehicle-treated mice.
Chronic administration of melatonin (2.5 or 5 mg/kg, i.p.) along with diazepam had no significant effect per se, but significantly reversed dose-dependent withdrawal-induced hyperlocomotor activity and severe anxiety.
Discussion
Benzodiazepines are one of the most widely used class of drugs in the management of anxiety, insomnia, muscle spasms, convulsions and alcohol withdrawal. However, their indiscriminate use has shown that these agents have a potential for producing physiological and psychological dependence.
All drugs including opiates and benzodiazepines when administered systemically lead to an increase in extra cellular levels of DA in mesolimbic pathway (22). Benzodiazepines produce drug dependence and withdrawal reactions on abrupt termination. These reactions involve both GABA / BZ receptor system and noradrenergic hyper-function in the brain. Apart from GABA, other neurotransmitter systems are also implicated in the psychopathology of benzodiazepine withdrawal. For example, diazepam withdrawal causes increased release of serotonin (5-HT) from the amygdala of rats, and this may contribute to the anxiogenic response (23). Exposure to anxiogenic drugs is also associated with increased, activity of the neurotransmitter dopamine (DA) in the nucleus accumbens (24).
The present study demonstrated that melatonin can attenuate the BZD abstinence syndrome. Melatonin administration alone for 21 days did not result in significant differences in loco motor behavior, which suggest the present attenuation effects upon BZD withdrawal were independent of motoricity.
Involvement of GABAergic and non-GABAergic mechanisms in withdrawal reactions are well documented. Melatonin significantly prevented the development of severe withdrawal to diazepam, the response being more pronounced at 24 h of withdrawal.
Melatonin is a safe drug having no apparent response per se on reward system. Concurrent administration of this drug may help in preventing the development of tolerance and dependence to benzodiazepines and other psychotropic drugs. The drug is also effective in combating withdrawal reactions to chronic administration of diazepam.
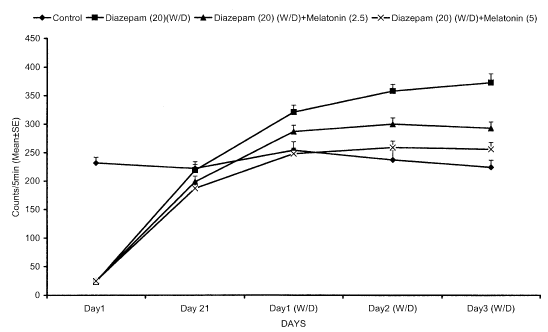
Fig. 1: Effect of co-administration of melatonin on ambulatory activity (measurement of loco motor activity) in diazepam-withdrawn mice. Values expressed mean ± SEM. P<0.05 as compared to diazepam treated group (for treatment schedule see text) (ANOVA followed by Dunnett's t-test).
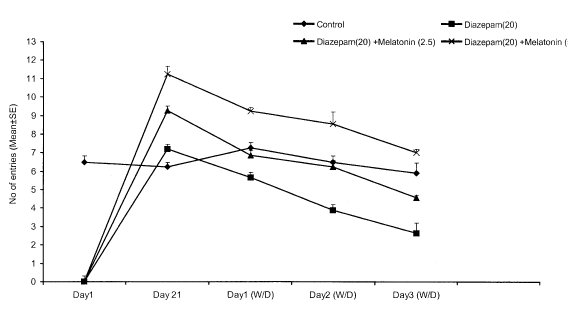
Fig. 2 (A): Effect of co-administration of melatonin on number of entries, in mirrored-chamber (measurement of anxiety) in diazepam withdrawn mice. Values expressed mean ± SEM. P<0.05 as compared to diazepam treated group (for treatment schedule see text) (ANOVA followed by Dunnett's t-test).
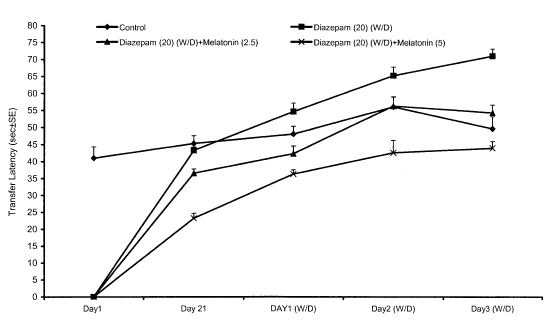
Fig. 2 (B).. Effect of co-administration of melatonin on transfer latency in mirrored-chamber (measurement of anxiety) in diazepam withdrawn mice. Values expressed mean ± SEM. P<0.05 as compared to diazepam treated group (for treatment schedule see text) (ANOVA followed by Dunnett's t-test).
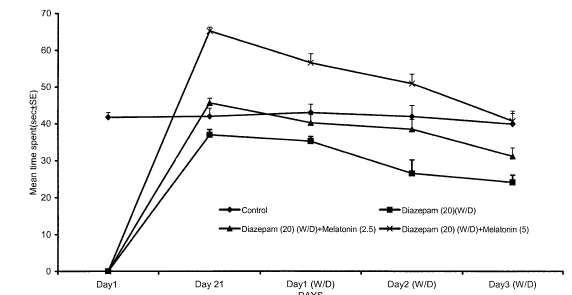
Fig. 2 (C). Effect of co-administration of melatonin on mean time spent in mirrored-chamber (measurement of anxiety) in diazepam withdrawn mice. Values expressed mean ± SEM. P<0.05 as compared to diazepam treated group (for treatment schedule see text) (ANOVA followed by Dunnett's t-test).
Melatonin (N-acetyl-5-methoxytryptamine), the chief indolamine produced by the pineal gland, has been shown to be an effective antioxidant and free radical scavenger (10–13). Melatonin, apart from direct free radical scavenging activity, indirectly enhances anti oxidative defense mechanisms by increasing the activities of several antioxidant enzymes and by stimulating the synthesis of another important intracellular antioxidant, glutathione (GSH) (14–15). Melatonin has been shown to exert an antioxidant effect on dopaminergic neurons, as well as dopaminergic modulating activities (17, 18). Melatonin, by acting through its own plasma membrane receptors, facilitates inhibitory GABAergic neurotransmitter functions, antagonizes 5-HT-2A/2C receptor-mediated behavioral responses and affects cholinergic neurotransmission in the nucleus accumbens (25–30). Besides these actions, at a cellular level melatonin suppresses nitric oxide synthase (NOS) activity via complex formation with cytoplasmic calmodulin in brain regions concerned with motor, autonomic and vegetative functions (31,32).
The present study provides evidence for the de-addiction potential of acute as well as chronic administration of melatonin against diazepam. The mechanism of action of reversal of diazepam tolerance and withdrawal-induced hyper loco motor activity and anxiety by melatonin, still remains to be explored.
Taken together, the present study shows that melatonin significantly reverses the development of diazepam tolerance and withdrawal-induced hyper loco motor activity and anxiety in mice.
References
1. Doble A, Martin IL. Multiple benzodiazepine receptors: no reason for anxiety. Trends Pharmacol Sci 1992;13(2):76–81.
2. Hutchinson MA, Smith PF, Darlington CL. The behavioral and neuronal effects of the chronic administration of benzodiazepine anxiolytic and hypnotic drugs. Prog Neurobiol 1996; 49: 73–97.
3. Schweizer E, Rickels K. Benzodiazepine dependence and withdrawal: a review of the syndrome and its clinical management. Acta Psychiatr Scand 1998; 393 (Suppl): 95–101.
4. Woods JH, Katz JL, Winger G. Benzodiazepines: use, abuse, and consequences. Pharmacol Rev 1992 Jun; 44(2): 151–347.
5. Rickels K, Demartinis N, Rynn R, Mando L. Pharmacologic strategies for discontinuing benzodiazepine treatment. J Clin Psychopharm 1999; 19:S12-S16.
6. Bateson AN. Basic pharmacologic mechanisms involved in benzodiazepine tolerance and withdrawal. Curr Pharm Des 2002; 8(1): 5–21.
7. Golombek DA, Povet P, Cardinali PD. Melatonin effects on behavior: possible mediation by the central GABAergic system. Neurosci Biobehav Rev 1996; 20(3): 403–12.
8. Pang SF, Wan Q, Brown GM. Melatonin receptors in the spinal cord. Biol Signals 1997; 6: 272–83.
9. Vanecek J. Cellular mechanisms of melatonin action. Physiol Rev 1998; 78: 687–721.
10. Reiter RJ, Tan DX, Acuna-Castroviejo D, Burkhardt S, et al. Melatonin: mechanisms and actions as an antioxidant. Current Top Bio-phys 2000; 24: 171.
11. Reiter RJ, Tan DX, Manchester LC, Qi W. Biochemical reactivity of melatonin with reactive oxygen species and nitrogen species. Cell Biochem Biophys 2001; 34: 237–56.
12. Bandyopadhyay D, Biswas K, Bandyopadhyay U, Reiter RJ, et al. Melatonin protects against stress-induced gastric lesions by scavenging hydroxyl radical. J Pineal Res 2000; 29: 143–51.
13. Bromme HJ, Morke W, Peschke E, Ebelt H, et al. Scavenging effect of melatonin on hydroxyl radicals generated by alloxan. J Pineal Res 2000; 29:201–8.
14. Reiter RJ, Tang L, Garcia JJ, Munoj-Hoyos A. Pharmacological actions of melatonin in oxygen radical physiology. Life Sci 1997; 60: 2255–71.
15. Urata Y, Honam S, Goto S, Tadoroki S, et al. Melatonin induces g-glutamylcysteine synthetase mediated by activator protein-1 in human vascular endothelial cells. Free Radic Biol Med 1999; 27(7–8): 838–47.
16. Reiter RJ. Pineal melatonin: cell biology of its synthesis and its physiological interactions. Endocr Rev 1991; 12: 151–80.
17. Kim YS, Joo WS, Jin BK, Cho YH, et al. Melatonin protects 6-OHDA-induced neuronal death of nigrostriatal dopaminergic system. Neuro Report 1998; 9: 2387–90.
18. Zisapel N, Egozi Y, Laudon M. Inhibition of dopamine release by melatonin: regional distribution in the rat brain. Brain Res 1982; 246(1): 161–3.
19. Kunchandy J, Kulkarni SK. Reversal by a2-agonists of diazepam withdrawal hyperactivity in rats. Psychopharmacology 1986; 90: 198–202.
20. Kulkarni SK, Sharma A. Reversal of diazepam withdrawal induced hyperactivity in mice by BR 16-A (Mentat), a herbal preparation. Ind J Expl Biology 1994; 32: 886–88.
21. Kulkarni SK, Reddy DS. Animal behavioral models for testing anti anxiety agents. Meth Find Exp Clin Pharmacol 1996; 18(3): 219–230.
22. DiChiara G, Imperato A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in mesolimbic system of freely moving rats. Proc Nat Acad Sci USA 1988; 85: 5274–5278.
23. Fernandes C, Andrews N, File SE. Diazepam withdrawal increases [3H]-5-HT release from rat amygdaloid slices. Pharmacol Biochem Behav 1994; 49(2): 359–62.
24. McCullough LD, Salamore JD. Anxiogenic drugs beta-CCE and FG 7142 increase extra cellular dopamine levels in nucleus accumbens. Psychopharmacology 1992; 109(3): 379–382.
25. Kopp C, Vogel E, Rettori MC, Delagrange P, et al. Antagonistic effects of S 22153, a new MT1 and MT2 receptor ligand, on the neophobia-reducing properties of melatonin in B alb/c mice. Pharmacol Biochem Behav 1999; 64: 131–6.
26. Wan Q, Man HY, Liu F, Braunton J, et al. Differential modulation of GABA A receptor function by Mel1a and Mel1b receptors. Nat Neurosci 1999; 2: 401–3.
27. Eison AS, Freeman RP Guss VB, Mullins UL, et al. Melatonin agonists modulate 5-HT 2A receptor mediated neurotransmission: Behavioral and biochemical studies in the rat. J Pharmacol Exp Ther 1995; 273: 304–8.
28. Drago F; Busa L, Benelli A, Bertolini A. Acute low doses of melatonin stimulate rat sex behaviour: the role of serotonin neurotransmission. Eur J Pharmacol 1999; 385:1–6.
29. Raghavendra V, Kulkarni SK. Melatonin reversal of DA I-induced hypophagia in rats; possible mechanism by suppressing 5-HT 2A receptor mediated activation of HPA axis. Brain Res 2000; 860:112–8.
30. Paredes D, Rada P, Bonilla E, Gonzalez LE, et al. Melatonin acts on the nucleus accumbens to increase acetylcholine release and modify the motor activity pattern of rats. Brain Res 1999; 850:14–20.
31. Pozo D, Reiter RJ, Calvo JR, Guerrero GM. Inhibition of cerebellar nitric oxide synthase and cyclic GMP production by melatonin via complex formation with calmodulin. J Cell Biochem 1997; 65: 430–42.
32. Bettahi I, Pozo D, Osuna C, Reiter RJ, et al. Melatonin reduces nitric oxide synthase activity in rat hypothalamus. J Pineal Res 1996; 20: 205–10.
(c) Annals of Neurosciences.All Rights Reserved