Annals of Neurosciences, Vol 14, No 2 (2007)
Annals of Neurosciences, Volume 14, Issue 2 (April), 2007
CYTOARCHITECTONIC PATTERN OF THE OLFACTORY SYSTEM, FOREBRAIN AND PITUITARY GLAND OF THE INDIAN MAJOR CARP, LABEO ROHITA
Address for Correspondance:
Professor VV Baile
Department of Zoology,
RTM Nagpur University Campus,
Nagpur 440 033 India
Tel: +91-712-2500-553; fax: +91-712-2563-356
Email:
Abstract
To study the organization and cytoarchitecture of the entire olfactory organ, forebrain and pituitary gland in the Indian major carp Labeo rohita, Kluver and Barrera and Aldehyde fuschsin staining techniques are used. In the olfactory system, important cell groups and nuclei identified are mitral cells, smaller sized granular cells and giant cells of nucleus terminalis. In the forebrain, important nucleus of larger sized neurons forming a cluster is located adjacent to the lateral forebrain bundle which is nucleus entopeduncularis in the telencephalon. Diencephalon is divided into preoptic area, hypothalamus, thalamus and epithalamus. In the preoptic area, both AF-positive and AF-negative neuronal groups are identified. Nucleus preopticus which is divided into preopticus pars paraventricularis and pars magnocellularis are AF-positive. In the preoptic area, nucleus of anterior commissure and nucleus preopticus periventricularis are AF-negative. On the ventrolateral side of the horizontal commissure (HC) in the preoptic area, nucleus of HC is localized. In the hypothalamus, behind the HC, tuberai area starts. In this area, nucleus hypothalamics is found on lateral side of the third ventricle. In the tuberai area near the infundibulum, nucleus lateralis tuberis is identified. In the caudal tuberal area, lateral side of the third ventricle shows nucleus recesses lateralis. Dorsal to the tuberal area, paraventricular organ is identified and nucleus inferior lobi is located on lateral side. Dorsal to the hypothalamus, thalamus shows dorsal and ventral thalamic nuclei. In the epithalamus, paired habenular ganglia are present dorsal to the thalamus. Pituitary is platybasic with short stalk and is forward in position.
Key words: Olfactory organ, Forebrain, Pituitary, Teleost, Indian major carp.
Introduction
Though teleostean brain conforms to the general vertebrate pattern, the unique feature is the absence of median eminence in teleosts and hypothalamic neurons directly innervating the pituitary cells. Teleostean hypothalomo-hypophysial system, therefore, offers a simple model to determine the anatomical relationship between the specific neurons, the bioactive substances present in them and the pituitary cell types on which they act. This type of brain structure has attracted the researchers over last few years. In several teleosts, olfactory bulb extends away from the telencephalon, thus increasing the length of olfactory peduncles or tracts. Such type of olfactory bulbs are known as pedunculated bulbs. They lie closely opposed to the peripheral olfactory epithelium (1,2). The cytoarchitectonic pattern of the forebrain of teleosts is studied in a number of species (Ictalurus punctatus:, Carassius auratus:, Ciarias batrachus:, Astyanax hubbsi:, Solea senegalensis:, Dicentrachus labrax) (1,3–7). Stereotaxic atlases for the forebrain nuclei of the goldfish Carassius auratus and the killifish: Fundulus heteroclitus are also available (3,8). Pituitary gland in teleosts is unique amongst the vertebrates in that it is directly innervated by the neurosecretory fibers (9).
Teleosts represent the most abundant group of bony fishes covering about 25,000 species, but the organization of olfactory system, forebrain and pituitary gland is studied in only a limited number of species (10). Furthermore, no information is available pertaining to the organization of the pituitary of any of the Indian major carps. Labeo rohita offers a good model to study cytoarchitecture of the olfactory system, forebrain and pituitary gland.
Materials and Methods
Adult males of Labeo rohita (n = 12) weighingl.5 kg to 2 kg and ranging in length between 30 to 40cms were collected from the natural habitat all around the City of Nagpur. They were brought to the laboratory in tin containers and acclimatized in small ponds. They were anaesthetized with 0.2% 2-phenoxy ethanol. Fishes were perfused transcardially with 750–850 ml ice cold phosphate buff ere saline (PBS, pH 7.45) followed by the same volume of ice-cold bouin's fixative. Olfactory organ with bulb and brain with pituitary gland were dissected out and post-fixed in the same fixative overnight. Olfactory organ and brain with pituitary gland were cut in transverse and sagittal planes at 10μm thickness. Alternate sections were stained with Nissl's stain and Aldehyde fuschsin to localize the neurosecretory material (11,12).
Kluver and Barrera technique
Sections of olfactory organ with bulb and brain with pituitary were deparaffinised in xylene and passed through different grades of alcohol bringing upto 95%. They were stained in 0.1% solution of Luxol Fast B1ue in 95% alcohol for 6 to 8 hours at 570C. Sections were rinsed in 95% alcohol to wash off the excess stain and brought down to distilled water through descending grades of alcohol, differentiated quickly in 0.1% Lithium carbonate solution and passed through several changes of 70% alcohol till there was a sharp contrast between the greenish blue white matter and colourless gray matter. Sections were washed thoroughly in distilled water and stained in 0.1% cresyl violet solution for 2 hours, then differentiated in several changes of 95% alcohol, dehydrated, cleared in xylene and mounted in DPX and coverslipped.
Aldehyde Fuschin (AF)
The deparaffinized sections of brain and pituitary were passed through descending grades of alcohol, brought to water and oxidized for 1 minute in 0.3% potassium permagnate solution acidified with sulphuric acid. They were decolorized quickly in 0.1% sodium metabisulphite. The sections were rinsed in distilled water and 70% alcohol, and then stained with AF, dehydrated in ascending grades of alcohol and mounted in DPX after clearing in xylene. Brain charts were prepared for the forebrain by plotting the sections from the series of transverse and sagittal sections. Each nuclear group in the forebrain was explored using cytoarchitectonic criteria.
Results
In the Indian major carp Labeo rohita, paired olfactory organs with bulb are located in the snout region. This organ is connected to the forebrain by the medial olfactory tract (MOT) and lateral olfactory tract (LOT).
Olfactory system
The olfactory organ has a central raphae, from which radiate lamellae on both the sides. These lamellae receive fascicles or fibers from its proximal end and extend into olfactory nerve (Fig-1). Olfactory nerve is short and caudally connected to the olfactory bulb. The olfactory bulb is divisible into four layers. Outer layer is olfactory nerve layer (ONL) which has axons of olfactory receptor neurons (ORN's). Below ONL, axons group together forming glomeruli called as Glomerular layer (GL). The glomeruli innervate the bigger sized neurons called mitral cells which form the Mitral cell layer (MCL). In the centre, densely packed small cells are present forming Granular cell layer (GCL) (Fig. 1, 4A, 4B).
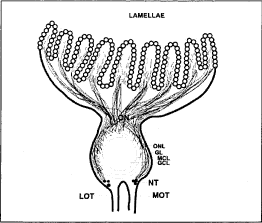
Fig. 1. Diagrammatic representation of the sagittal section of the olfactory system of L. rohita showing, GCL, granular cell layer; GL, glomerular layer; MCL, mitral cell layer; MOT, medial olfactory tract; ON, olfactory nerve; ONL, olfactory nerve layer.
The fibers from the olfactory nerve penetrate the olfactory bulb peripherally and innervate profusely forming glomerular structure over the larger sized mitral cells which show intense Niss'l staining (Fig. 4A). Some smaller cells found in GCL also show intense Niss'l staining. The cells which are dorsal to olfactory bulb situated at the base of olfactory bulb are intensely stained and these are giant cells of nervus terminalis (NT) (Fig. 4B).
Forebrain
Forebrain in Labeo rohita is divided into telencephalon and diencephalon.
Telencephalon
Telencephalon is divisible into two main regions i.e. area dorsalis telencephali and area ventralis telencephali. Its organization is shown in figure-2,3. Area dorsalis telencephali is subdivided into pars medialis (Dm), pars centralis (Dc), pars lateralis dorsal (Dld) and pars lateralis ventral (Dlv). Small sized cells located near the medial region of dorsal telencephalon is Dm which show moderate Niss'l staining. The neurons in the area Dc are few in number and show intense Niss'l staining. Cells in the area Did are rounded in shape and are moderately stained, while neurons in the area Dlv are spindle shaped, small in size and moderately stained. Area ventralis telencephali is broad, divided into pars ventralis (Vv), pars dorsalis (Vd) and pars lateralis (VI). Neurons in the Vv are sparsely arranged, smaller in size and exhibit moderate Niss'l staining. Cells of the Vd are bigger as compared to the cells of Vv, compactly distributed and show intense Niss'l staining. In the area VI, few larger sized cells showing intense staining are observed. The medial and lateral forebrain bundles are present in vetrolateral position of the telencephalon. Telencephalic lobes are connected with each other by Anterior commissure and Commissure of Goldstein which are located dorsal to the preoptic recess. These two commissures are parallel to each other (Fig. 4C).
Adjacent to the forebrain bundle, few bigger sized neurons are seen forming a cluster which is nucleus entopeduncularis (NE) (Fig. 4D). It shows intense Niss'l staining (Fig. 4D). However these neurons are AF negative.
Diencephalon
In Labeo rohita, diencephalon includes four regions- preoptic area, hypothalamus, thalamus and epithalamus.
In the preoptic area, hypothalamus forms a major part of ventral diencephalon. Preoptic area is the portion of prosencephala wall that covers the preoptic recess of third ventricle. It is covered anteriorly by Anterior commissure and commissure of Goldstein, dorsally by telencephalon and ventrally by the optic chiasma. Preoptic area contains both AF positive and AF negative neuronal groups. The neurons of the Nucleus preopticus (NPO) are AF positive and other nuclei in the preoptic area are AF negative.
Neuronal complex of the preoptic area is paired in paraventricular region of the preoptic recess which is AF negative. Nucleus of anterior commissure (NAC) is one of the nuclei of preoptic areawhich is AF-negative. It is located near the ventral margin of anterior commissure and dorsal margin of the commissure of Goldstein on lateral side of the preoptic recess. Neurons of NAC are spindle shaped and show weak Niss'l staining. Another AF negative nucleus is nucleus preopticus periventricularis (NPP). It is present close to the preoptic recess on both the sides. NPP is divided into nucleus preopticus periventricularis dorsalis (NPPd) and ventralis (NPPv) (Fig. 2). Cells of NPPv are small, aggregated and show intense Niss'l staining. Neurons of NPPd area are comparatively less in number and show moderate Niss'l staining.
In Labeo rohita, neurons of NPO are present on either side of the preoptic recess. On the basis of size of these neurons, NPO is divisible into NPO pars paraventricularis (NPOp) and NPO pars magnocellularis (NPOm). Neurons of NPOp are located above the optic chiasma and around the third ventricle, NPOm neurons are larger in size as compared to the NPOp and are located dorsally around the third ventricle (Fig. 2, 3). Neurons of NPOp and NPOm show intense Niss'l staining (Fig. 4E). These are AF positive and show accumulation of neurosecretory material in the cytoplasm of NPOp and NPOm. They have long axons which collectively form the hypothalamo-hypophysial tract which innervates the pituitary gland.
Suprachiasmatic nucleus (SCN)
On either side of the third ventricle is located the SCN (Fig. 2). It has small spindle shaped neurons which show moderate Niss'l staining.
Nucleus of horizontal commissure (NHC)
Horizontal commissure is present in the post optic area. It has paired nuclei that cover the horizontal commissure on ventrolateral side (Fig. 2). The nuclei are rounded and show intense Niss'l staining.
Nuclear groups of tuberal area
Behind the Horizontal commissure, tuberai area starts. It consists of neurons located on either side of the third ventricle and its lateral recess. In the tuberai area, lateral side of the third ventricle shows three prominent nuclei i.e nucleus hypothalamics ventralis (Nhv), nucleus hypothalamicus dorsalis (Nhd) and nucleus hypothalamicus medialis (Nhm) (Fig. 2, 3). Nhv consists of comparatively small cells and, shows moderate Niss'l staining. Above Nhv, round shaped cells of Nhm show weak Niss'l staining. Dorsal to the Nhm, spindle or round shaped cells of Nhd show moderate staining. Nucleus lateralis tuberis (NLT) is an important nuclear group of the tuberai area located near the infundibulum (Fig. 2,3). It is present ventral to the horizontal commissure and extends along the base of hypothalamus. NLT is divisible on the basis of position and size of the neurons into NLT pars lateralis (NLT1) and NLT pars medialis (NLTm). NLT1 neurons are located laterally, large in size and rounded in shape showing intense Niss'l staining (Fig. 4F). NLTm neurons are located near the third ventricle and comparatively smaller in size than NLTl neurons. NLTm cells are few in number and show moderate Niss'l staining.
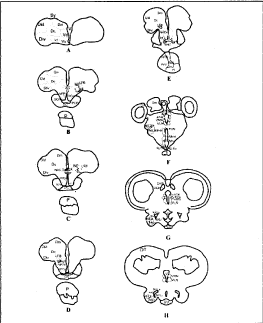
Fig. 2. Diagrammatic representation of the transverse section of the forebrain of L. rohita showing, AC, anterior commissure; ATN, anterior thalamic nucleus; CG, commissure of Goldstein; Dc, area dorsalis telencephali pars centralis; DHN, dorsal habenular nucleus; Did, area dorsalis telencephali pars lateralis dorsal; Dlv, pars lateralis ventral; Dm, pars medialis; HC, Horizontal commissure; LFB, Lateral forebrain bundle; NE, Nucleus entopeduncularis; NH, Neurohypophysis; NHC, Nucleus horizontal commissure; Nhd, nucleus hypothalamicus dorsalis; Nhm, nucleus hypothalamicus medialis; Nhv, nucleus hypothalamicus ventralis; NILd, nucleus inferior lobi pars dorsalis; NILv, nucleus inferior lobi pars ventralis; NPOm, Nucleus preopticus pars magnocelularis; NPOp, nucleus preopticus pars parvocellularis; NPPd, nucleus preopticus periventricularis dorsal; NPPv, nucleus preopticus periventricularis ventral; NRL,-nucleus recesses lateralis; NRLi, nucleus recesses lateralis inferior; NRLs, nucleus recesses lateralis superior; OC, optic chiasma; OT, optic tectum; P, pituitary gland; PI, pars intermedia; POA, preoptic area; POR, preoptic recesses; PPD, proximal pars distalis; PTN, posterior thalamic nucleus; PVO, paraventricular organ; RPD, rostral pars distalis; SCN, suprachiasmatic nucleus; SV succus vasculosus; V, ventricle; VHN, ventral hypothalamic nucleus; VLH, ventrolateral thalamic nucleus; VMN, ventromedial thalamic nucleus.
Caudal to the tuberai area, lateral side of the third ventricle give rise to the lateral recesses. Compactly arranged cells on the periphery of the lateral recesses are nucleus recesses lateralis (NRL) (Fig. 2, 3). This nucleus is again sub grouped into superior and inferior groups on the basis of their position i.e. nucleus recesses lateralis superior (NRLs) located on the dorsal side of the lateral recess and nucleus recesses lateralis inferior (NRLi) present on the ventral side of the lateral recess. The NRLs consists of large number of cells and show moderate Niss'l staining. Cells of NRLi are smaller in size and compactly arranged as compared to the NRLs cells. They show intense Niss'l staining.
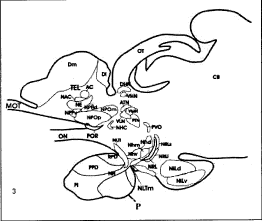
Fig. 3. Diagrammatic representation of the saggital section of forebrain and pituitary gland showing, AC, anterior commissure; ATN, anterior thalamic nucleus; CB, cerebellum; Dm, area dorsalis telencephali pars medialis; DHN, dorsal habenular nucleus; Dl, pars lateralis; MOT, medial olfactory tract; NE, Nucleus entopeduncularis; NH, Neurohypophysis; NHC, Nucleus horizontal commissure; Nhd, nucleus hypothalamicus dorsalis; Nhm, nucleus hypothalamicus medialis; Nhv, nucleus hypothalamicus ventralis; NILd, nucleus inferior lobi pars dorsalis; NILv, nucleus inferior lobi pars ventralis; NPOm, Nucleus preopticus pars magnocellularis; NPOp, nucleus preopticus pars parvocellularis; NPPd, nucleus preopticus periventricularis dorsal; NPPv, nucleus preopticus periventricularis ventral; NRL, nucleus recesses lateralis; NRLi, nucleus recesses lateralis inferior; NRLs, nucleus recesses lateralis superior; OC, optic chiasma; ON, optic nerve; OT, optic tectum; P, pituitary gland; PI, pars intermedia; POR, preoptic recesses; PPD, proximal pars distalis; PTN, posterior thalamic nucleus; PVO, paraventricular organ; RPD, rostral pars distalis; SCN, suprachiasmatic nucleus;TEL, telencephalon VHN, ventral hypothalamic nucleus; VLH, ventrolateral thalamic nucleus; VMN, ventromedial thalamic nucleus.
Dorsal to the tuberal area and paraventricular in position is Paraventricular organ (PVO) which demarcates the dorsal margin of hypothalamus. The cells of PVO are in contact with CSF (Fig. 2,3).
Nuclear groups of inferior lobes
Nucleus inferior lobi is located on the lateral side which is divisible into nucleus inferior lobi pars dorsalis (NILd) and nucleus inferior lobi pars ventralis (NILv) (Fig.2, 3). The cells of the NILd are spindle shaped and show intense Niss'l staining. Neurons of the NILv are comparably smaller in size, round shaped that show moderate Niss'l staining.
Pituitary gland
Pituitary of L. rohita is piatybasic type. It is attached to the hypothalamus by a short stalk and is forward in position. Axons of the hypothalamic nuclei penetrate the neurohypophysis of pituitary gland. The pituitary gland is divided into adenohypophysis and neurohypophysis. Adenohypophysis is subdivided into rostral pars distalis (RPD), proximal pars distalis (PPD) and pars intermedia (PI). AF positive cells are seen in the PPD region (Fig.4H). These cells are large in size and show the accumulation of secretary material.
Thalamus
Thalamus is made up of two divisions: dorsal and ventral thalamus (Fig. 2,3). In the dorsal thalamus, cells of anterior thalamic nucleus (ATN) are rounded and spindle shaped, abundant in number and show intense staining of Niss'l. Neurons in the posterior thalamic nucleus (PTN) are comparably less in number, spindle or rounded in shape and show moderate Niss'l staining. Ventromedial thalamic nucleus (VMN) and ventrolateral thalamic nucleus (VLN) are parts of the ventral thalamus. The neurons of the VMN are spindle shaped and show intense Niss'l staining. The cells in the VLN are comparably smaller is size, more in number and show moderate Niss'l staining.
Epithalamus
In the epithalamus, a pair of habenular ganglion are present dorsal to the thalamus and two habenular ganglia are connected by a habenular commissure. Each habenular ganglion has two nuclei i.e. Dorsal habenular nucleus (DHN), and ventral habenular nucleus (VHN). The cells in the VHN are rounded and show intense Niss'l staining (Fig.4G). In the DHN, cells are comparably less in number and show moderate Niss'l staining (Fig.4G).
Discussion
In Labeo rohita, olfactory bulb is of pedunculated type, connected to the telencephalon. Such type of olfactory organ is also found in other members of family Cyprinidae and Acanthidae (1). Olfactory bulb comprises of four layers from superficial to the deep in L. rohita. The fibers of the ORNs extend caudally over the olfactory nerve and penetrate into the bulb from anterior side and spread along the periphery of the bulb in some teleosts which are also seen in L. rohita (2,13,14). In the olfactory bulb of L. rohita, at the base of olfactory tract, the ganglion cells of the nervus terminate have been identified. Smaller cluster of the deeply stained ganglion cells along vertromedial surface of the nervus terminalis is also noted in Ictalurus punctatus (1). Telencephalon in L. rohita consists of two hemispheres connected to each other by Anterior commissure (AC) and Commissure of Goldstein (GC) and is basically divided into area dorsalis telencephali and area ventralis telencephali. Other teleosts also show the same pattern of division (5, 6, 15). The area dorsalis telencephali is subdivided into Dm, Dc, Did, Dlv according to review by Meek and Nieuwenhuys and Rodriguez-Gomez et al. (6,16). In number of other teleosts, area ventralis telencephali is subdivided into Vv, Vd, VI and NE depending on the area which they occupy and same is observed in L. rohita (3,6,8,16).
Diencephalon is considered to be the most complex region in the brain of the teleosts (7). It is the neuronal integration centre in the goldfish for gustatory information, reproduction and vision (17). Recently, using cytoarchetectonic criteria, epithalmus, thalamus, hypothalamus and preoptic area are included as the parts of the diencephalon in Dicentrachus labrax (17). This type of division of diencephalon is also used in the cytoarchitectural analysis to study the organization of the neuronal groups in the brain of teleosts (3, 5, 6, 7, 8, 17). In L. rohita, same pattern of division is observed. Preoptic area is the part of rostralmost diencephalic subdivision (16). A dense aggregation of nuclei around the third ventricle is an important landmark to localize the preoptic area. This area has been subdivided into magnocellular, parvocellular, preoptic nucleus and suprachiasmatic nucleus (Carassius auratus:, Haplochromis:) (17,18). In the present study, AF positive small NPO cells referred as pars parvocellularis and large cells called as pars magnocellularis are observed as shown in some of the teleosts, situated at the level of third ventricle (19, 20). Other workers employed the term magnocellularis for AF positive and parvocellularis for AF negative cell groups. In the preoptic area, AF negative smaller cell groups are divided into Nucleus preopticus periventricularis dorsalis (NPPd) and ventralis (NPPv) which are comparable to that of Clarias batrachus (4). The nucleus of anterior commissure in L. rohita is important because of its association with the anterior commissure. This nucleus is also described in other fish and reptiles (4, 21). SCN nucleus in L. rohita is an aggregation of small cells which is also reported in other fish, amphibians, reptiles and mammals (22, 23, 24). This nucleus receives retinal projections (25). NLT has been extensively studied in several teleosts (4, 19). In some teleosts, NLT neurons are reported to be AF positive (26,27). In L. rohita also, they are AF positive. Functionally, the role of NLT in control of gonadotropic cells is well documented in number of teleosts (28,29).
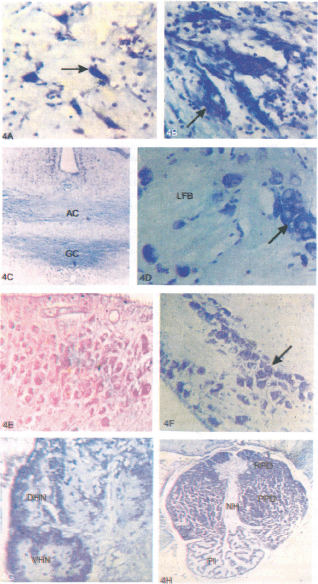
Fig. 4. Sagittal and transverse sections of olfactory bulb and brain of L. rohita. A) Olfactory bulb showing larger sized mitral cells with intense Niss'l staining (arrow) and B) bigger sized neurons of nervus terminalis (NT) (arrow). C) Telencephalon showing anterior commissure (AC) and Commissure of Goldstein (CG)and D) bigger size neurons adjacent to the lateral forebrain bundle (LFB) in nucleus entopenducularis (NE) (arrow) with intense Niss'l staining. E) Diencephalon showing intense staining of Aldehyde Fuschin in nucleus preopticus pars magnocellularis (NPOm) and F) tuberai area showing bigger sized neurons located laterally showing intense Niss'l staining in nucleus lateralis tuberis pars lateralis (NLTl) (arrow). G) Epithalamus showing habenular ganglion with dorsal habenular nucleus (DHN) and ventral habenular nucleus (VHN). H) Transverse section of pituitary gland showing Neurohypophysis (NH), rostral pars distalis (RPD), proximal pars distalis (PPD) and pars intermedia(PI). Magnifiaction: (A,B),400X; (C, E, F, G), 160X; (D), 500X; (H), 25X.
In L. rohita, on the lateral side of the third ventricle, the anterior tuberal area consists of Nhm, Nhd and Nhv. The Nhm and Nhd are comparable with the nucleus periventricularis posterior of Corydora palitus but in other vertebrates it is described as a dorsal hypothalamic nucleus in a similar position (17,18,30).
In caudal part of the tuberal area, on the lateral side of the third ventrical, hypothalamus gives extensions called as the lateral recesses: lateralis superior and inferior in L. rohita. In salmonids, catecholamine producing cells of nucleus recesses lateralis (NRL) and NRL in Clarias batrachus seem to be the similar ones. The nucleus paraventricular organ (NPVO) associated with paraventricular organ (PVO), shown in Clarias batrachus is comparable with the one found in L. rohita (4).
In L. rohita, inferior lobe in hypothalamus has two prominent nuclei, nucleus inferior lobi pars dorsalis (NILd) and nucleus inferior lobi pars ventralis (NILv) but earlier workers have shown only one nucleus i.e. nucleus diffuse lobi inferioris (3,31). Same findings are noted in Clarias batrachus (4). In L. rohita, thalamus is divided into dorsal and ventral portions. In the dorsal division, there is anterior thalamic nucleus (ATN) and posterior thalamic nucleus (PTN), while in the ventral thalamus, two nuclei are identified i.e. the ventrolateral thalamic nucleus (VLN) and ventromedial thalamic nucleus (VMN), but in Dicentrachus labrax, dorsal thalamus is formed by three nuclei: the ATN, PTN and the central posterior thalamic nucleus. Ventral thalamus is made up of nucleus of thalamic ementia, the ventrolateral thalamic nucleus, ventromedial thalamic nucleus and the intermediate thalamic nucleus (7, 17, 18). Such type of division however, is not clearly marked in L. rohita.
Epithalamus region in L. rohita consist of habenular ganglion and the habenular commissure. Habenular ganglion consist of the dorsal and ventral nuclei, as reported in other teleosts (7,16, 17).
Pituitary gland in L. rohita is platybasic and is divided into adenohypophysis and neurohypophysis. Adenohypophysis is subdivided into rostral pars distalis (RPD), proximal pars distalis (PPD) and pars intermedia (PI). Similar pattern is also reported in other teleosts (9,32).
As teleost is a big group of fishes showing enormous diversity, it is important to uncover the underlying pattern of brain to determine the anatomical and functional correlation of its various parts. Teleosts are further unique in vertebrates in that there is direct innervation of hypothlamic nuclei in adenohypophysis. This study will thus be helpful to find out the correlation of brain and pituitary gland in the Indian major carps.
References
1. Bass AH. Olfactory bulb efferents in the channel catfish, Ictalurus punctatus. J Morph 1981; 169: 91–111.
2. Baby SM, Ueck M, Prasada Rao PD. Gonadotropin-releasing hormone-immunoreactive neurons and associated nicotinamide adenine nucleotide phosphate-diaphorase-positive neurons in the brain of teleosts, Rhodeus amarus. Gen Comp Endocrinol 2000; 120: 44–54.
3. Peter RE, Gill VE. A stereotaxic atlas and technique for forebrain nuclei of the goldfish, Carrassius auratus. J Comp Neurol 1975; 159: 69–102.
4. Ramakrishna NS, Subhedar N. Cytoarchitectonic pattern of hypothlamus in the catfish, Clarias batrachus (Linn.). J Hirn Forsch 1991; 32: 289–308.
5. Riedel G. The forebrain of the blind cave fish Astynanax hubbs (Characidae). Brain Behav Evol 1997; 41: 20–38.
6. Rodriguez-Gomez FJ, Rendon-Unceta MC, Sarasquete C, Munoz-Cueto JA. Localization of tyrosine hydroxylase-immunoreactivity in the brain of the Senegalese sole, Solea senegalensis. J Chem Neuroanat 2000; 19: 17–32.
7. Cerda-Reverter JM, Zanui S, Munoz-Cueto JA. Cytoarchitectonic study of the brain of a perciformis species, the sea bass (Dicentrarchus labrax), II The diencephalon J Morphology 2001; 247: 229–51.
8. Peter RE, Macey MJ, Gill VE. A stereotaxic atlas and technique for forebrain nuclei of the killifish, Fundulus heteroclitus. J Comp Neurol 1975; 159:103–28.
9. Peter RE, Yu KL, Marchant TA, Rosenblum PM. Direct neural regulation of the teleost adenohypophysis. J Exp Zool Suppl 1990; 4:84–89.
10. Nelson GJ. Origin and diversification of teleostean fishes, Ann NY Acad Sci 1969; 167: 18–30.
11. Kluver H, Barrera E. A method for the combined staining of cells and fibers in the neurons system. J Neuropathal Exp Neurol 1953; 12: 400–403.
12. Gabe M. Surquilquies applications de la coloration per le fuchsineparaldehyde. Bull Micr Appi 1953; 3:153–62.
13. Khan FA, Saha SG, Sarkar S, Subhedar N. β-endorphin-like immunoreactivity in the forebrain and pituitary of teleosts Clarias bafrachus (Linn.) Gen Comp Endorcrinol 1999; 113: 290–301.
14. Singru PS, Sakharkar, AJ, Subhedar, NK. Neuronal nitric oxide synthase in the olfactory system of an adult teleost fish, Oreochromis mossambicus. Brain Res 2003; 977: 157–68.
15. Bass AH. Telencephalic effernts in the channel catfish, Ictalurus punctatus: Projections to the olfactory bulb and optic tectum. Brain Behav Evol 1981; 19:1–16.
16. Meek J, Nieuwenhuys R. Holosteans and teleosts In: (Eds) R Nieuwenhuys, HJ Ten Donkelaar, C Nicholson. Central Nervous System of Vertebrates 1998; 759–937.
17. Braford M, Northcutt R. Organization of the diencephalon and pretectum of the rayfinned fishes In: (Eds.) R Davis, R Northcutt Fish Neurobiology Vol. II: Higher brain areas and functions: University of Michigan Press, 1983.
18. Fernald R, Shelton L. The organization of the diencephalon and the pretectum in the cichlid fish, Haplochromis burtoni. J Comp Neurol 1985; 238: 202–17.
19. Prasada Rao PD, Betole UK. Changes in the hypothalamo-hypophysial complex after gonadectomy in the catfish, Ciarias batrachus (Linn.). Zool Beitr 1973; 19:319–33.
20. Peter RE, Fryer JN. Endocrine functions of the hypothalamus of Actinopterygii. In: (Eds) RE Davis, RG Northcutt Fish Neurobiology. Vol. II, The University of Michigan Press, 1983; 165–201.
21. Prasada Rao PD, Subhedar N, Raju PD. Cytoarchitectonic pattern of the hypothalamus in the cobra, Naja naja. Cell Tissue Res 1981; 217: 503–29.
22. Neary TJ, Northcutt RG. Nuclear organization of bull frog diencephalon. J Comp Neurol 1983; 23:262–78.
23. Prasada Rao PD, Subhedar N. Cytoarchetectonic study of the hypothalamus of the lizzard, Calotes versicolor. Cell Tissue Res 1977; 180: 63–85.
24. Arimura A, Findley A. Hypothalamic map for the regulation of gonadotropin released based mainly on data obtained in the rat. Res Reroduct 1971; 3:1.
25. Springer AD, Mednick AS. Selective innervations of the goldfish suprachiasmatic nucleus by ventral retinal ganglion cell axons. Brain Res 1984; 323:293–96.
26. Belsare DK, Belsare SG, Murthy PSR. Hypothalamo-hypophysial neurosecretory system in normal and hypophysectomised catfish, Clarias batrachus (Linn.) Acta Zool 1970; 51: 219–2
27. Sathyanesan AG. Hypothalamo-hypophysial neurosecretory system of the freshwater teleost, Catla catla (Ham.) Zool Beitr 1972; 11: 163–78.
28. Sathyanesan AG. The nucleus preopticus of the fresh water catfish, Clarias batrachus (L.) Zool Beitr.l970; 16:247–55.
29. Billard R, Peter RE. Gonadotropin release after implantation of anti-estrogen in the pituitary and hypothalamus of goldfish, Carassius auratus. Gen Comp Endocrinol 1977; 32: 213–20.
30. Millar RN. The diencephalic cell masses of the teleosts, Corydora paliatus. J Comp Neurol 1940; 73: 345–78.
31. Billard R, Peter RE. A stereotaxic atlas and technique for nuclei of the diencephalon of rainbow trout (Salmo gardeneri). Reprod Nutr Dev 1982; 22:1–25.
32. Ball JN, Baker BI. The pituitary gland: Anatomy and histophysiology In: (Eds) WS Hoar, DJ Randal. Fish Physiology. 1969; 1–110.
(c) Annals of Neurosciences.All Rights Reserved