Annals of Neurosciences, Vol 13, No 4 (2006)
Annals of Neurosciences, Volume 13, Issue 4 (October), 2006
ENDOCRINOLOGICAL STUDY WITH THE BRAIN OF NON MULBERRY SILKWORM ANTHERAEA ASSAMA USING IN VITRO TECHNIQUE
Corresponding Author
Dr. BG Unni
Bio-Technology Department,
Regional Research Laboratory,
Council of Scentific And Industrial Research,
Jorhat, Assam, India
E-Mail:
Abstract
The in vitro study with the brain of non mulberry silkworm showed that the brains collected from the second to fourth day of fifth instar larvae were suitable to survive in culture media as represented by the assay of aldolase activity. The soluble protein content in the brains kept in vitro, for 18 hours at dark was found to be lower than the same observed for the fresh brains. The native PAGE analysis of the soluble protein extract of the fresh and incubated brain collected from Antheraea assama revealed three bands for the former and two for the later. The band having the lowest mobility was not detected in the protein extract of the incubated brain. It suggested probable hydrolysis or migration of that particular protein during incubation. Absence of the soluble protein in the media used, emphasized the possibility of hydrolysis of protein due to action of protease as certain reports justified the need of soluble protein not only for the growth of insect cell in vitro but also to protect the cultured cells from the action of protease. However, as reported earlier for other insects, the migration of peptides from the brain tö the media was also confirmed with the help of native PAGE analysis of the media (after incubating 40 brains for 18 hours at 30°C at dark).
Key words: Antheraea assama, Soluble Protein, Aldolase, Native PAGE
Introduction
Brain is the most important part of the endocrine system releasing biomolecules, peptides, etc. which on the other hand governs many functions, including silk protein biosynthesis. Use of cell culture techniques for better understanding of insect endocrinology have been reported (1–5). In vitro culture of the brain have been achieved in numerous systems. However, its application to silkworm particularly to Antheraea assama is not known. Studies relating to detection, isolation and identification of brain released factors, together with determining their probable role in silk protein biosynthesis will certainly help in better understanding of the endocrinolpgy of the less studied silkworm Antheraea assama. Attempts involving in vitro isolation of such factors from semi-domesticated muga silkworm exposing their probable positive effect on silk protein biosynthesis or inducing hibernation of muga silkworm will be beneficial to the farmers of sericulture. Present study was basically an attempt to standardize the conditions required for in vitro culture of brains collected from silkworm, Antheraea assama.
Material and Method
Muga silkworm and its rearing for in vitro study:
The disease free layings (DFLs) of Antheraea assama were received from Central Muga Research and Training Institute, Lahdoigarh, Jorhat. The eggs were disinfected by dipping in formaline solution (2%) for 10 minutes. When the first instar larva emerged, within 12 hours they were transferred to the host plants (Persea bombycina) covered with the net. Required number ten of larvae of desired instar (fifth instar) were collected for both in vivo and in vitro study.
Culture of silk worm brain in the media:
The brains collected from ether anesthetized muga silkworm (fifth instar) were cultured in the Grace's media (GIBCO, Grand Island, NY, USA) and the insect tissue culture media (Hi-media, Bombay) according to standard procedure (6). Dissected brains were washed in the another lot of the same media and finally transferred to the sterilized media (containing phenyl thio urea) kept in autoclaved micro dishes. Gentamycin was added to the Grace's media before the transfer of the brain @ 0.5%. Prior to transfer of the brain, the micro dishes were ringed with sigma cote (sigma, USA) and washed with double distilled water. After transfer of the brain, the covered micro dishes were incubated in a incubator for 18 hours at 30°C in dark. After incubation period, the brains were removed and the media checked for infections under the microscope and pooled and stored at 4°C for further analysis. The incubated intact brains were transferred to a micro homogenizer with definite volume (300ml) of sterilized distilled water, homogenized in ice and then the extract was subjected to the following analysis in two phases.
First phase:
- Total soluble protein: The soluble protein was extracted by precipitating the protein of the homogenate (150/μl) by adding double the volume of 10% tri-chloro-acetic acid (TCA), keeping in ice for 1 hour, followed by centrifuging at 6000rpm for 10 minutes at room temperature and then dissolving the precipitate in definite volume (l00ul) of 0.1M NaOH. The total soluble protein was determined by standard procedure (7).
- Glucose and pyruvate: After centrifugation, the supernatant was used for estimation of glucose and pyruvate (8,9).
- Aldolase: The aldolase activity was assayed in the water homogenate (150/μl) (10).
The above mentioned parameters were determined in controlled (un-incubated) brain also and the data from both the sets (incubated and un-incubated) were compared.
Second phase:
Based on the results of the first phase of this experiment, the second phase was planned. In this phase, the brains from only the fifth instar early stage (4th day old) larvae were isolated. After incubation period, the incubated larval brain and media (after separation of the brain from it), both were subjected to native Poly Acryl amide Gel Electrophoresis (PAGE) (11) to determine probable presence and nature of small peptides released by the brain during incubation time.
Preparation of samples for PAGE:
- The brains were removed from the clear media after the end of the incubation period, washed in sterilized distilled water, transferred to a micro homogenizer, definite volume (150μl) of sterilized distilled water was added, homogenized, to this solution double the volume of 10% TCA was added, kept at 0°C for 1hour to precipitate the protein. Then the precipitate was collected, dissolved in aliquot of distilled water (50μl) and double the volume of sample buffer was added and brought to the room temperature.
- The plain media (where brain was not incubated) and the entire amount of the test media (where brain was incubated) after separating the brains from it, were collected, then lyophilized completely to dryness. This is dissolved directly in definite volume (150μl) of sample buffer (pH 6.8) and brought to the room temperature.
Result
The comparison of both the in vivo and the in vitro (in Grace's media) aldolase activity, soluble protein, glucose and pyruvate content in the brain of the muga silkworm (Antheraea assama) are presented in the <
| Stages | Total activity (units) |
Soluble protein (×10−3 mg) |
Specific activity (units/mg protein) |
Pyruvate (×10−5 mM/brain) |
Glucose (×10−5 mM/brain) |
|---|---|---|---|---|---|
| Vth I,0d(F) | 0.137 | 13.43 | 10.20 | 5.00 | 10.3 |
| Vth I,0d(V) | 0.000 | 7.68 | 0.000 | 7.50 | 8.8 |
| Vth I, 2d(F) | 0.099 | 11.47 | 8.63 | 0.00 | 12.0 |
| Vth 1, 2d(V) | 0.113 | 7.24 | 15.61 | 8.00 | 12.6 |
| Vth I, 4d(F) | 0.597 | 17.55 | 34.02 | 0.00 | 11.0 |
| Vth I, 4d(V) | 0.656 | 10.97 | 59.59 | 16.67 | 0.7 |
Data are averages of three independent samples.
One unit of aldolase activity (total or specific) is defined as change in optical density by 1 unit at 540 nm/hour/brain or/mg protein.
Aldolase activity in the brain:
Except on the day of moulting, the aldolase total activity of brain in vitro were observed to be higher on the second day and fourth day of fifth instar than the respective in vivo activity. Moreover, the variations of specific activity were observed to be similar for fresh and incubated brains. For fresh brains, the lowest and the highest level of specific activity were detected on second day of fifth instar and fourth day of fifth instar, respectively. For the brains after incubation, the same was observed to be the lowest on the day of moulting and the highest on fourth day of fifth instar.
Soluble protein content of the brain:
It was observed that the soluble protein content was lower in the brains kept in vitro than the same observed for the brain kept in vivo. The difference in soluble protein between fresh and incubated brain ranged from 4.23 μg/brain observed for second day of fifth instar to 6.58 μg/brain for fourth day of fifth instar.
Glucose content of the brain:
It was observed that on the day of moulting, the glucose content in the fresh brain was lower than the same present in the second day of fifth instar or the fourth day of fifth instar larval brain. For Antheraea assama, the glucose content of the fresh brain was observed to be 10.3 x 10−5 mM/brain on the day of moulting, whereas the same were recorded to be 12.0 x 10−5 mM/brain and ll.0 x 10−5 mM/brain for second day and fourth day of fifth instar larval brain, respectively.
The glucose content of the brains of Antheraea assama after incubation was observed to be the highest (12.6 x 10−5 mM/brain) at second day of fifth instar stage whereas the same was observed to be the lowest (0.7 x 10−5 mM/brain) for the brains of fourth day of fifth instar stage.
Pyruvate content of the brain:
The pyruvate was not detected in the fresh brains of Antheraea assama, collected from both the second day and fourth day of fifth instar stages. For the brains after incubation, the lowest and the highest level of pyruvate were identified on the fifth instar fourth day stage and on the day of moulting, respectively.
Qualitative differences in the soluble protein content of fresh and incubated brain:
The native PAGE analysis (
The native PAGE analysis (
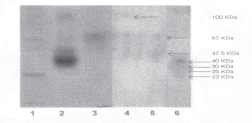
Fig. 1: Non SDS-PAGE analysis of soluble protein extract collected from fresh brain, incubated brain and media where brains of Antheraea assama were incubated.
- Standard, 24 KDa;
- Standard, 45 KDa;
- Standard, 66 KDa;
- Soluble protein extract (Fresh brain);
- Soluble protein extract (Brain kept in vitro);
- Media (after incubation of brain);
Discussion
The present study was conducted to standardize the condition for in vitro survivability of the brain of the muga silkworm Antheraea assama. Analysis of the data has shown that on the day of moulting, the brain is not suitable to survive in vitro condition which might be due to the presence of initial low level of glucose in the brain (10.3 x 10−5 mM/brain). Though glucose was present in the media, the brain might require certain initial level of the glucose in the cells of the brain at the time of inoculation to the media. Brains of fifth instar second day and fourth day larvae of Antheraea assama were more capable form to survive in vitro as at these two larval stages, the aldolase total activity per brain was found to be sufficiently higher in vitro condition than the respective in vivo activity. The glucose was well utilised by many insect tissues when cultured in media (12). 90% utilisation of glucose present in the Grace's media by the cells of Antheraea eucalypti grown for seven days was observed (13). After depletion of glucose reserve of the brain, the cell may utilise glucose of the media through diffusion. Diffusion of trehalose and branch chained amino acids from media to tissue kept in vitro were also reported (14,15).
One of the remarkable observation was the lower soluble protein content of the brains which were kept in vitro. This might be due to migration of some peptide to the media during incubation. In vitro secretion of a small peptide, called 'allatinhibin' from the larval brain of the Manduca Sexta has been reported (6). It has been reported about secretion of a form of ion transport peptide (ITP) in an insect cell expression system(5). Migration of peptides to the media during incubation of the brains was also confirmed from the present study as native PAGE analysis of the media showed presence of four bands on silver staining. Among these four bands, the intensity of peptide having the lowest mobility was maximum.
There was also possibility of degradation of certain peptide of the brains kept in vitro. In a study, foetal bovine serum (FBS) protein in insect tissue culture medium was used (16). It contained eight protein fractions of which about 40–50% was represented by fetuin (an alpha – glycoglobulin) (17,18). Disc electrophoresis of fresh tissue culture medium and medium from seven day old cultures revealed that 13% of the protein at the albumin region and 67% of protein at the fetuin region had disappeared (16). From this study, they suggested that specific proteins or substances bound to them were required and metabolised by Aedes aegypti cells cultured in vitro. The cockroach cell line (Epa) also exhibited a requirement for proteins (19). They also suggested that the protein might be beneficial because they function as anti-protease and thus protect the cultured cells from proteolytic enzymes. It is worth mentioning that both the media which were used for the present in vitro study were lacking the protein in their composition. Therefore, the possibility of utilisation (for the activities of proteolytic enzymes present in the brains) of protein present in the cells of the brain during incubation for metabolic purposes can not be eliminated. In the present investigation the native PAGE analysis of the soluble protein extract prepared from equal number of brains (of the same stage), both fresh and kept in vitrorevealed the presence of two similar bands on silver staining (
The study was successful in not only establishing the stage of the larvae of Antheraea assama from which the brains should be collected for in vitro study but also to identify the qualitative and quantitative changes in the soluble protein content of the incubated brain from that of the fresh brain. Therefore, isolation and further identification of functional properties of brain released factors or peptides would be very useful in understanding the mechanism of juvenile hormone biosynthesis or silk protein biosynthesis or any other physiological functions (including hibernation) in insects. This information may have some important functions in planning an improvement in the area of muga sericulture.
Acknowledgement:
The authors are thankful to Dr. PG Rao, Director, Regional Research Laboratory, Jorhat for permission to publish this research article.
References
1. Chinzei V. Induction of histolysis by ecdysterone in vitro: Breakdown of anterior silk gland of silkworm Bombyx mori. (Lepidoptera: Bombydae). Appl Entomol Zool 1975; 10:136.
2. Hatt PJ, Moriniere M, Oberlander H, Porcheron P. Roles for insulin and ecdysteroids in differentiation of an insect cell line of epidermal origin. In Vitro Cell Dev Biol Animal. 1994; 30: 717–20.
3. Levine RB, Weeks JC. Cell culture approaches to understanding the actions of steroid hormones on the insect nervous system. Dev Neurosci 1996; 18: 73–86.
4. Oberlander H, Siibacek DL. Mode of actions of insect growth regulators in Lepidopteran tissue culture. Pesticide Sci 1998; 54 : 300–02.
5. Pfeifer TA, Hegedus D, Wang YJ, et al. Analysis of an insect neuro peptide, Schistocersa gregaria ion transport peptide (ITP), expressed in insect cell systems. Arch Insect Biochem Physiol. 1999; 42: 24–25 .
6. Unni BG, Barrera P, Muszynska-pytel M, Bhaskaran G, Dahm KH. Characterisation of allatinhibin, a neurohormone of Manduca sexta. Arch Insect Biochem Physiol. 1993; 24:173.
7. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the folin – phenol reagent. J Biol Chem. 1951; 193:265.
8. Thompson RH. Determination of glucose by glucose oxidase method. Clinical Chemistry Acta. 1966; 13: 133.
9. Fiedman TE. Determination of alpha keto acids. In: Methods of Enzymology Colowick SP, Kaplan NO Eds., New York: Academic Press; 1963:414.
10. Sibley JA, Lehninger AL. Determination of aldolase in animal tissues. J Biol Chem 1949; 177:859.
11. Laemmli Cleavage of structural proteins during the assembly of the head of .bacteriophage T4. Nature 1970;227:680–85.
12. Wyatt GR, Kalf GF. The chemistry of insect haemolymph. J Gen Physiol 1957; 40:833.
13. Grace TDC. Establishment of a line of mosquito (Aedes aegypti L) cells grown in vitro. Nature 1966; 211:366–67.
14. Jabbar TUA, Mohamed UVK. In vitro studies on trehalose diffusion and the occurrence of trehalase in the gut of the larvae of Orthaya exvinacea Hampson. (Pyralidae: Lepidoptera) Comp Physiol Ecol, 1993; 18:136–39.
15. Brundle PA, Baker FC, Tsai LW, Reuter CC, Schooley DA. sources of propionate for the biosynthesis of ethyl branched insect juvenile hormone: Role of isoleucine and valine. Proceed Natl Acad Sci USA 1987; 7906–10.
16. Kuno G. Studies of growth promoting proteins in fetal bovine serum using Aedes aegypti cells cultured in vitro. PhD Dissertation, The Ohio State University, Ohio, USA. 1970.
17. DiDomizio G, Minoccheri F. Sieroproteine fetaliu ed evoluzione postnatate del ferogramma sierico bovino. Arch Vet Ital 1964; 15: 9–20.
18. Spiro RC. Studies on fetuin, a glycoprotein of fetal serum. Isolation, chemical composition and physicochemical properties. J Biol Chem 1960;235: 2860–69.
19. Landureau JC, Steinbuch M. Cyanocobalamine as a support of the in vitro cell growth promoting activity of serum proteins. Experimentia 1969; 25: 1078–79.
(c) Annals of Neurosciences.All Rights Reserved