Annals of Neurosciences, Vol 17, No 3 (2010)
Annals of Neurosciences, Volume 17, Number 3, July 2010
Biochemical and cytogenetic analysis of brain tissues in different grades of glioma patients
KEY WORDS
Glioma
Antioxidants
Enzymes
Chromosomal aberrations
Biochemical profile in glioma
Corresponding Author
Vanisree Arambakkam Janardhanam, Ph.D
Tel.: +91-44-26493652
Fax: +91-44-22352494
E-mail: vanielango@gmail.com
ABSTRACT
Background: Glioma, a neoplasm of neuroglial cells, is the most common type of brain tumor, constituting more than 50% of all brain tumors. Purpose: This report summarizes the current knowledge regarding the clinical utility of biochemical enzyme markers for both diagnostic and therapeutic purposes in different grades of glioma. Methods: Sixty patients with different grades of glioma include glioblastoma multiforme (n=20), Anaplastic astrocytoma (n=10). Oligodendroglioma (n=10). Ependymoma (n=10), Pilocytic astrocytoma (n=10) and patients with benign lesions (n=5) served as controls. Activity of antioxidants and marker enzymes were assayed in all grades followed by karyotyping. Results: Activities of antioxidant enzymes reduced significantly (p<0.05) compared to controls except CAT, GST that showed marked increase (p<0.05) in accordance with malignancy. CK, Na-K+ATPases, 5’-Nucleotidases showed marked increase in grade IV. Similarly, Mg2-ATPase, Ca2+ATPases showed significant increase in grade III. Conclusion: The clinical importance for classification and treatment of glioma is governed by biochemical enzyme markers. The study of enzymes supported by related chromosomal changes is anticipated to provide better appreciation of biological properties in different grades of glioma.
doi: 10.5214/ans.0972-7531.1017305
Introduction
Glioma, a neoplasm of neuroglial cells, is the most common type of brain tumor, constituting more than 50% of all brain tumors. It accounts for about 2% of the malignant tumors in adults and is a leading cause of central nervous system (CNS) tumor related death.1 The most common forms of primary brain tumors are the gliomas, including the highly malignant glioblastoma multiforme which have a high rate of re-incidence after surgical resection.2 There prevalence is 23% among all brain tumors in Chennai.
Conventionally, brain tumors are diagnosed by imaging. In some cases, the information obtained from imaging may be inadequate to determine the type and / or grade of brain tumors a patient has. To further characterize brain tumors, the tumor may be biopsied to determine the type and grade of the tumor which includes histopathological examination however it involves a certain degree of subjectivity. Unfortunately, conventional histopathology techniques often do not provide definitive guidance whether low grade tumors will progress to high grade.3 In addition to diagnostic procedures, the assessment of biochemical enzymes becomes essential that are subject to regulation by various signalling pathways which are prominent in particular grade of glioma determining their biological behaviourand resistance pattern.
Numerous studies revealed that the most common somatic chromosomal changes in malignant gliomas are complete or partial loss of chromosome.4 Such loss or deletion represents not only the tumor suppressor gene but also marker enzymes involved in various signaling mechanism including cell cycle control. Thus, it would be a valuable study to assess the prognosis of such abnormalities of brain tumor for determining the most appropriate course of treatment for a glioma patient.
Based on the above concept, the enzymatic profile including the antioxidant status was determined in brain tumor patients which plays an important role in biochemical metabolism and also in signaling network in different grades of glioma. The metabolic data was correlated with cytogenetic data giving the relative numbers of chromosomes or chromosomal segments carrying the gene encoding for each enzyme.
Methods
Subjects
After the informed consent of all patients and approval of the ethical committee, the tumor samples were collected from different grades of glioma patients (n=60) and patients with benign lesions (n=5) during surgery at the Institute of Neurology and Neurosurgery, MMC, Chennai. Fresh tissue sections of (n=20) Glioblastoma Multiforme (GBMs), (n=10) low grade glioma, (n=10) ependymoma (ED), (n=10) anaplastic astrocytomas (AA), (n=10) oligodendrogliomas (OA) samples were obtained. The pathological classification of tumor tissues was carried out and the stage of each glioma patients was determined according to the WHO classification.5 GBM patients were 25 -72 years old (mean = 53 years, n = 20), OA patients were 20-68 years old (mean = 45 years, n = 10), AA were 21 -60 years old (mean = 34 years, n = 10), grade II ED patients were 23-62 years old (mean = 50 years, n = 10), grade I pilocytic astrocytoma patients were 18-52 years old (mean = 34 years, n = 10) and BL (mean=22, n=5).
Biochemical Analysis
Excised fresh brain tumor tissues were taken and used for enzyme analysis described below. Total protein was estimated by the method of Lowry et al. Superoxide dismutase was assayed by the method of Marklund et al.7 and was expressed as U/min/mg protein. The method of Beer and Sizer8 was adopted for the assay of catalase (CAT). The activity was expressed as H2O2 decomposed/min/mg protein. The method of Rotruck et al was adopted for the assay of glutathione peroxidase (GPx) and enzyme activity was expressed in nmoles of GSH oxidised/min/mg protein. Activities of glutathione-s-transferase (GST) was assayed by the method Habig et al.10 and expressed in nmoles of CDNB conjugated/min/mg protein. Glutathione reductase, (GR) was assayed by the method Stall GE et al11 and activity expressed as u/mg protein. 5’-nucleotidase (5’- ND), was estimated by the method Sandra et al12 and activity expressed as Mmoles/min/mg protein. The methods of Horrocks13 was adopted for the assay Lactate dehydrogenise (LDH) and activity expressed as/imoles of pyruvate liberated/min/mg protein. The activity of Aspartate Transaminase in tissue sample was assayed by the method of King et al14 and enzyme activity was expressed in μmoles of pyruvate liberated/min/mg protein. The Alkaline phosphatases (ALP) activity was assayed by the method of Green et al15 and activity was expressed in micromoles of p-nitrophenol liberated/min/mg protein. The activity of creatine kinase was assayed by the method Norman Hall et al16 and enzyme activity was expressed in IU /mg protein. Isolation of plasma membrane was done by the method Aileen et al.17 Activity of mg2+ATPases was assayed by the method Aileen et al and units expressed as μmoles of phosphorous liberated/mg protein/min. Na+, K+ATPase activity were assayed according to Tsakiris et al18 and units were expressed as /jmoles of phosphorous liberated/mg protein/min. Activity of Ca-ATPases assayed by the method Junkosato et al19 and units were expressed as Ca2+ ATPases in μmoles of phosphorous liberated/min/mg protein.
Karyotypic analysis
The tissue specimens were collected and transported in 10ml 0.5% sodium citrate + 0.5μg colcemid/ml (total time in solution: 1 hr). They were disaggregated by scraping and cutting in a petridish and filtered through a 100μ nylon filter. After incubation in Hank’s Balanced salt solution, Colcemid was added (2μg/ml Hank’s BSS) for 30min. Hypotonic treatment was done with a mixture of 6ml fetal calf serum and 24ml 0.052 M KCI. Fixation was performed with acetic acid: methanol 3:7. The G-band pattern was carried out using three-week-old slides treated with 0.01% trypsin in phosphate buffer (pH 7.2 ) for 5 min, rinsed in the same buffer, added with 0.1% D-glucose, and stained with 5% Giemsa in pH 6.8 phosphate buffer for 30s.
Statistical analysis
Data was analyzed using SPSS for Windows V.10. One way analysis of variance (ANOVA) was performed to find out the significance of variations between three groups followed by Student’s t-test and Spearman’s correlation test was conducted for correlation analyses.
Results
Activities of biochemical enzymes among different grades of glioma varied with the most significant difference existing between the grades. Comparisons of the antioxidants marker enzymes activity of the various histological subtypes are shown in Table land 2. Significant increased level of total protein were observed in glioma samples compared to control and it was progressively increased with a higher grade of malignancy (Table 2). The activities of transaminase (AST) and phosphatase (ALP) were increased compared to control, the increased activity AST, ALP were significant in grade IV (p<0.001) and grade III (p<0.05) when compared to control.
Tumor tissues showed a reduction in activities of SOD, GPX, and GR in comparison to benign tissue. The increased activities of enzyme CAT and GST showed significant changes in high grades compared to other grades and benign lesions. There was also a significant difference between the low grade and high grade tumor tissue (Table 1).
A statistically, significant (p<0.05) increase of LDH, CK, and ATPases were found in all grades compared to benign. The activity was (4-8 fold) higher in the tumors than in the control group (Table 2).
Table 1: Comparison of activity of antioxidant enzymes between the grades
| Enzyme activity | Grade I | Grade II | Grade III | Grade IV | BL |
| SOD units/min/mg protein |
8.10±1.2@ | 6.28±0.82* | 5.43±0.36* | 4.863±0.45*• | 9.01±0.9 |
| CAT μmoles H2O2 consumed/min/mg protein |
3.0±0.41@ | 3.8±0.28@ | 5.94±0.8*†† | 6.12±0.72*†@ | 3.36±0.28 |
| GST Mmoles/min/mg of protein |
132±12*1 | 38±22.2*@ | 178±22.1**† | 192±20*†• | 120.9±16 |
| GR u/mg protein |
31.16±3.4@ | 30±3.1 | 28±2.9 | 22.5±2.54*† | 30.94±28.2 |
| GPX nmoles /min/mg protein |
42.2±6.5@ | 38.1 ±4.0* | 32.08±4.3* | 29.11 ±3.89*†† | 44.32±5.2 |
*P<0.05 -compared to BL
• P<0.05 -compared between grade III & IV
††P<0.01, †P<0.05 - compared between low & high grades
@-not significant
On comparisons between different grades of glioma CK, Glu6PDH, 5-nucleotides, Na-k+ ATPases and GST showed marked (P<0.05) difference in their activities in GBM when compared to other grades. Similarly, Ca2+ATPases, Mg2+ATPases (P<0.01) show increased activity in grade III compared to other grades. The observed enzyme profile was compared with chromosomal aberrations in benign and in different grades of glioma.
Table 2: Comparison of activity of marker enzymes in different grades of glioma
| Enzyme activity | Grade1 | Grade II | Grade III | Grade IV | BL |
| CK IU /mg protein | 98.2±9.5* | 110± 10.2*† | 126±12**•† | 140±12.9**•† | 90.8 ± 0.06 |
| 5-NUCLEOTIDASE mmoles/min/mg protein |
0.78±0.05@ | 1.24±0.08** | 2.89±0.02*† | 3.92±0.54*•† | 0.69±0.051 |
| AST μmoles of pyruvate liberated/min/mg protein |
8.72±0.8@ | 8.95±1.0@ | 9.01 ±0.99*@ | 9.23±1.52*@ | 8.4±0.72 |
| ALP micromoles of p-nitrophenol iberated/ min/mg protein |
4.12±0.24@ | 4.95±0.82@ | 4.9±0.9@ | 5.02±0.35**@ | 4.08±0.33 |
| LDH μmoles of pyruvate liberated/min/mg protein |
20.45±3.14@ | 28.5±2.2@ | 34.12±4.01†** | 38±5.8**@† | 19.24±2.2 |
| Na+/K+ ATPases in μmoles . of phosphorous liberated/ mg protein/min |
15±1.02 | 22.0±2.8*† | 28.11±4.2*† | 35.6±2.24*•† | 13.29±1.4 |
| Ca2+ ATPases in μmoles of phosphorous liberated/ mg protein/min. |
4.82±0.4@ | 5.10±0.35*@ | 6.8±0.5*@† | 5.5±0.32*@ | 3.7±0.32 |
| Mg2+ ATPases in μmoles of phosphorous liberated/ mg protein/min |
0.12±0.01* | 0.18±0.01*@ | 0.34±0.02*†† | 0.21 ±0.01 *†@ | 0.06±0.01 |
**P<0.01,*P<0.05- compared to BL
•P<0.05, ††P<0.01- compared between grade III & IV
†P<0.05 not significant - compared between low and high grades.
Karyotypic analysis
Table 3, shows the loss and amplification of chromosome observed and the most common loss found to be located on the long arm of chromosome 6 (6q,11p13, 14q24.1, 11q13), chromosome 10, while gains of chromosomes on 7,8q, 15,12,19, 20 and 21 were highly correlated with one another in all grades. These loss and gain of this chromosomal region well correlated with the genes specific for antioxidant and marker enzymes which is presented in graph I (a,b).
|
|
Graph: (a) Comparison between the loss of chromosomes and loss |
Graph: (b) Comparison between the gain of chromosomes and |
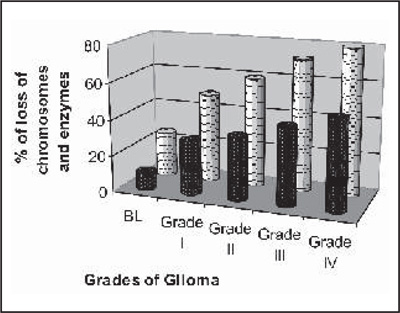
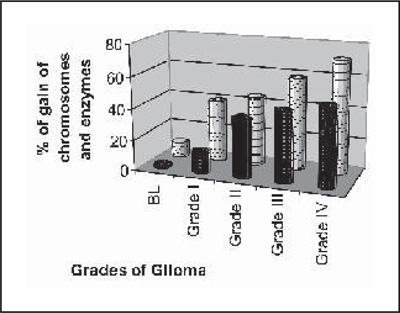
Discussion
In the clinical practice of oncology, understanding the prognosis of a given tumor is of paramount importance to patients and clinician alike. Analyzing the activity of marker and antioxidant enzymes may provide vital link in diagnosing different grades of tumor. The molecular pathogenesis of glioma includes both loss and gain of proteins responsible for cell proliferation and multiplication.20,21 Protein levels of glioma patients were found to be increased significantly when compared to normal as confirmed by Rongcai Jiang et al22 suggesting that the active protein biosynthesis occurring in the tumor cells, facilitates the tumor progression, such as degradation of the extra cellular matrix to promote tumor invasion. A significantly increased level of protein in GBM (p<0.05) compared to other grades suggest that complex pattern of alterations likely to involve multiple cellular pathways and molecular regulatory systems in the aggressive tumor cells. Thus, a comprehensive survey of protein expression and functionally important modification is particularly useful in providing insight into the working of altered pathways during disease progression, i.e from low to high grade (GBM). Many studies illustrate the well known observation that cell transformation alters cell responsiveness to oxidative stress.23 In neoplastic transformation, antioxidant enzyme activities have shown different patterns and they are highly variable in tissues and blood of patients with different types of cancer.24,25,26
Table 3: Karyotype analysis shows common aberrations in different grades of giloma
| GRADES | KARYOTYPE |
| Low grade | 46,XX,+7,-10*43,XX,-6,+7,-10,13,-16,del(18)*44,XY,-10,-1 (p?),*47,XX,+7,- 10,+2*44,XY,+7,-9,-10,-18,del(1 )(q32q43),t(6;16)*46.XX,+7,-10* 47,xx,+12,*45xy,-16,*45,xx,-22,*47,XX,+21,*45XX,-6(P?) |
| Anaplastic astrocytoma |
46,t(X;9)(q26-8;p12)Y,t(7;10)(q31;q21),*45,XY,-9*48,XY,+Y,+7,+12,-14 *44,xx,+,x,46,xx,-1(p),-22*47,xy,+21*44,xx,+19,*44,xx,-1(P),-22* 48,XY,+Y,+7,+ 12,-14,del(15)(q12q21 ),*44,xx,-18,-22 |
| Oligodendroglioma | 43,XY;+7,-10,-13,-19,-21,-(1)(p21-2),del(6)(q13-14), del(9)(p13), *47,xy,+19,45,xx- p(p?(q?),*45,xx,-6(p?)*46,xx,del(11),q(21),*41,xx,del(1)(p?),-10,-14,-16,-18,- 22,*45,x,-x,47,xx,+7 |
| Ependymoma | 46,t(X;5)(q36-8;p10)Y*49,XX,+3,+7,+9,+11* 49,XY,+7,+9(q),+20,*47,XX,+5,+6,*43,XY,-1,-2,-10,*45,XX,-1(p),-6(p), *45,XY,-1(p),*44,XX,+2,-22,+9,*45,XX,-15(q),*43,XX,-1(p),-13(q),-14(q),- 22(q),*46,XY,+3,-6 |
| GBM | 45,XX,del(1)(p?),-10,-14,-16,-18,-22*45,-x,-y*46,xxdel(11)(del22)* 45,xy(17)(q?),*47,xy,(+91)*46,xy,del(21)(q21),*45,xx,+x*46,xx,del(1)(p?),41,xx,d el,-19,22*47,xy,(1)(p?)*47,xy,+21*45,xy,-19,-22*46,xy,del(10)(p?)(q?),* 49,XY,+1,+7,+19,t(15;17)(q22;q25),*46,XX,+7,-10,*50,XX,+1,+1,+ 18,+18,0,der(7)t(7;?)(p22;?)+der(7)t(7;?)(p22;?)/26X,+1,+7,15,+18,-t-der(9)t(9;15) (q 11,q 11),der(11)t(11;?)(q24;?),*45,XX,del(1p?)(6p?) |
Our observation in tumor tissue showed a reduction in the activities of antioxidant enzymes (SOD, GPX, GR) in ascending order from low grade to high grade (Table 2) as supported by Pu et al27 and Rolando et al.28 Reduction in the activities of antioxidant in these tumors suggests that the tumor cells are able to survive and proliferate despite the higher than-average level of reactive oxygen species they exhibit. This is generally taken as a support to the role of ROS in cell proliferation. In contrast with the other antioxidant enzymes CAT(P<0.01), GST(P<0.05) were found to be distinctly increased in high grade compared to benign type and within different grades of glioma suggesting the high expression of GST in tumor cells and the presence of this protein in tumor cell nuclei are associated with clinically more aggressive gliomas. Further GST is associated with failure of cancer chemotherapy and in poor prognosis.29 Binding of GST-Pi to JNK reduces the intrinsic activity of JNK and decreases activation of JNK caused by cellular stresses and decreased apoptosis. Elevated catalase activity in high grade was reported by Pameeka30 stating that primary antioxidant enzyme, catalase, is elevated constitutively in gliomas compared to normal astrocytes as also observed in the current study. Increased activity of phosphatases is a constant feature in neoplastic transformation31 and accordingly, in the study, an increase in the activities of transaminases (ALT) and phosphatases (ALP) in different grades were found when compared to control.
Increased activities of enzymes (LDH, CK, 5’-ND, ATPases in different grades of glioma have been shown in Table I. Increased LDH suggests enhanced LDH activities in brain tumor. Catecholamine, through cAMP, have been shown to increase the activity of LDH in glioma.32 This elevation could damage the tumor cells in the brain. We observed elevated activity of CK which has central role in regeneration of ATP in the brain as tumor cells require high energy for cell division and invasion. 5’-N which is an endonuclease, whose chief function is to degrade the purine and pyrimidine nucleotides.33 Here, elevated 5’,N activity with respect to higher grade of glial malignancy may be due to its involvement in cell proliferation and progression. This process appears to depend upon the production of adenosine, a proliferative factor and removal of AMP, a toxic molecule for tumor cells.
In the present study, there was a considerable increase in all the membrane bound ATPases in the glioma tissues compared to control. Ion gradients are present across cell membranes and produce voltage across membrane to drive proper signaling functions across neuron. Ion gradients also aid in ATP generation and for uptake of neurotransmitters. It is identified that normally glial cells have a higher K+ permeability than all the other neurons and Na+ current is constantly elevated in malignant Glioma but not in normal dividing cells.34 In our report, when compared between the grades of Na/K-ATPase activity it was found to be highly elevated (p<0.05) in GBM than the other grades. Thus Ion pumps play pivotal role in cancer cell migration in general and in glioblastomas (GBMs) in particular. However, the specific role of Na/K-ATPase (the sodium pump), has remained unexplored in GBM.35 Similarly Mg2+-ATPases was found to be marked increase (P<0.01) in grade III compared to other grades. Na+-K+-ATPase was overwhelmed by Mg2+-ATPase in the astrocytoma which was striking. Thus, the excessive Mg2+-ATPase activity of the astrocytoma might be of significance in dedifferentiation or neoplastic transformation. The possibility that the high ATPases activity in the plasma membranes of the human tumors was also closely correlated with tumorigenicity should be seriously considered.
Chromosomal aberrations
Various structural chromosomal aberrations were identified, among which aberrations involving chromosome 1,6,10,16,21 were frequently seen in different grades of glioma support with report Maria et al.36 Human malignant gliomas are frequently associated with loss of gonosomes and chromosomes 13,17, and 22. Their progression from anaplastic glioma to glioblastoma is marked by additional loss of chromosome 10. Further, structural and numerical aberrations of chromosome 7 were frequently found.37 A significant association was observed between status of antioxidant enzymes and chromosomal deletions in different grades of glioma. Interestingly, we found reduced activities of SOD, GR, GPX by virtue of their significant deletions and loss of chromosome 6 that encodes these genes (6q,11p13,14q24.1,11q13), while gain in chromosome 11,12,15,21 that represent genes corresponding to catalase, LDH, GST isoenzymes, CK which shows increase in their activities in glioma tissue. The activity of 5’Nucleotidase, ATPases appeared to be unrelated to the relative number of chromosomes 6,19,16, respectively, that carry their encoding genes.
Certain genes have a consistently higher prevalence of allelic loss than other genes, including 19p and 16q, but appreciable changes were observed on many chromosomes in each case of glioma. This diversity, heterogeneity, and variation in specificity of genetic abnormalities suggest that there are multiple pathways for development of varied grades of glioma, many of which could hypothetically be triggered or influenced by epigenetic events, including oxidative damage.
The observed activities of enzymes and related chromosomal aberrations shows their involvement in metabolic energy and signalling pathways that allowed a characterization of energy demands of gliomas in different grades compared to control. Though the interpretation of the observed data is difficult for obtaining a vivid picture, the present study might stimulate further investigations for a better management of glioma.
Competing interests - None, Source of Funding - None
Received Date : 6 April 2010; Revised Date : 10 May 2010
Accepted Date : 29 May 2010
References
1. Phatak P, Selvi SK, Divya T, et al. Alterations in tumour suppressor gene p53 in human glioma from Indian patients. J. Biosci 2002; 27:673-678.
2. Deepak Kumar G, Mattei Tobias A, Ashok Kumar M, et al. Future of genetherapies in high grade gliomas. Cancer Therapy 2005; 3:555-564.
3. Robert G, Whitmore MD, Jaroslaw Krejza MD, et al.Prediction of oligodendroglial tumor subtype and grade using perfusion weighted magnetic resonance imaging. J Neurosurg 2007; 107:600-609.
4. Keshava R, Jothi R, Gope ML, et al.Functional modulation of the P53 gene and its protein in human brain tumors. Annals of Neurosciences 2008;15:75-86.
5. Louis DN, Holland EC, Cairncross JG. Glioma Classification A Molecular Reappraisal. Am J Pathol 2001;159:779-786.
6.[??] Lowry OH, Rosenbrough NJ, Farr AL, et al.Protein measurement with folin’s phenol reagent. J Biol Chem 1951 ;193:265-276.
7. Marklund S, Marklund G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur J Biochem 1974;47:469-474.
8. Saraf MK, Prabhakar S, Anand A. Neuroprotective effect of Bacopa Mommiera on ischemia induced brain injury. Pharmacol Biochem Behav (2010) doi 10.1016/J.P.bb2010.07-017.
9.[??] Rotruck JT, Pope AL, Ganther HE. Selenium: Biochemical role as a component of glutathione peroxidase purification and assay. Sciencel 973;179: 589-590.
10. Habig WH, Pabst MJ, Fleischner G, et al. The dentity of glutathione S transferase B with ligandin, a major binding protein of liver. Proc Natl Acad Sci U S A1974;71:3879-3882.
11. Stall GE, Visser J, Veeger C. Purification and properties of glutathione reductase of human erythrocytes. Biochimica et Biophysica Acta 1969;185:39-48.
12. Sandra I, Patrizia B, Lo Bello M, et al. Insulin-dependent release of 5•-nucleotidase and alkaline phosphatase from liver plasma membranes. Bioscience Reports 1992;12:101 -108.
13. Horrocks JE, King J, Waind APB, et al. Lactate dehydrogenase activity in the diagnosis of malignant effusions. J Clin Pathol 1962;15:57-61.
14. King J. Practical clinical enzymology. London: D. Van. Nostrand Company 1965;83-93.
15. Green JB, Oldewurtel H, O’doherty DS, et al. Cerebrospinal fluid glutamic oxalacetic transaminase activity in neurologicdisease. Neurology 1957;7:313-322.
16. Norman HI, Paul A, Marlene D. Mitochondrial creatine kinase. Physical and kinetic properties of the purified enzyme from beef heart. Biochemistry 1979;18:1745-1751.
17. Aileen F, Jose F, Nathan O. Isolation and Characterization of Plasma membranes from Transplantable Human Astrocytoma, Oat Cell Carcinoma, and Melanom. Cancer research 1981;41:4031 -4038.
18. Tsakiris S, Deliconstantinos G. Influence of phosphatidylserine on (Na+ K+)- stimulated ATPase and acetylcholinesterase activities of dog brain synaptosomal plasma membranes. Biochem J 1984; 22:301-307.
19. Junko S, Ritwko M, Hisashi T. The possibility that Ca’+-ATPase from the plasma Membrane rich fraction of bovine parotid gland is Ecto-Cam+-dependent nucleoside triphosphatase, int. J. Biochem 1994;26:905-910.
20. Ahmed Rasheed BK, Wiltshire Rodney N, Bigner Sandra H, et al. Molecular pathogenesis of malignant gliomas. Curr Opin Oncol 1999;11:162-167.
21. Van de K. Molecular pathogenesis of astrocytoma and glioblastoma multiforme. Acta neurochirurgica, 1997;39:589-599.
22. Rongcai J, Cristian M, llya S, et al. Pathway alterations during glioma progression revealed by reverse phase protein lysate arrays. Proteomics 2006;6:964-971.
23. Cross CE, Halliwell B, Edward TB. Oxygen radicals and human disease. Ann Int Med 1987;107:526.
24. Ahmed Ml, Fayed ST, Hossein H, et al. Lipid peroxidation and antioxidant status in human cervical carcinoma. Dis Markers 1999; 15:283-291.
25. Hristozov D Gadjeva V, Vlaykova T, et al. Evaluation of oxidative stress in patients with cancer. Arch Physiol Biochem 2001 ;109:331 -336.
26. Abiaka C, Al-w-awadi F, Al-sayer H, et al. Activities of erythrocyte antioxidant enzymes in cancer patients. J Clin Lab Anal 2002; 16:167-171.
27. Pu Y Lan J, Shan SB, et al. Study of the antioxidant enzymes in human brain tumors. J Neuro Oncol 1996; 29:121.
28. Rolando DM, Warren M, Robert A. SOD, catalase and glutathione peroxidase in experimental and human brain tumors. Oxyradicals and theirscavenger systems 1983;2:83-28.
29. Ali-osman F, Brunner JM, Kutluk TM, et al. Prognostic significance of glutathione S-transferase pi expression and subcellular localization in human gliomas. Clin Cancer Res 1997;3:2253-2261.
30. Pameeka S.Weiling Zhao ,Mike Robbins. Knocking down catalase leads to increased ROS production and proliferation in glioma cells. Proc Amer Assoc Cancer Res 2006;33:123-124.
31. Filmus JE, Podhajcer OL, Mareso E, et al. Acid phosphatases in human breast cancer tissue. Cancer 1984;53:301-305.
32. Miles MF, Hung P, Jungmann RA. CyclicAMP regulation of lactate dehydrogenase. Quantitation of lactate dehydrogenase M-subunit messenger RNA in isoproterenol-and N6,02’-dibutyryl cyclic AMP-stimulated rat C6 glioma cells by hybridization analysis using a cloned cDNA probe. J Biol Chem 1981 ;256:12545-52
33. Marcia R, Winkguido L, Elizandra B, et al. Altered extracellular ATP, ADP and AMP catabolism in glioma cell lines. Cancer Letters 2003; 198-95.
34. Bakhrom K, Jiazeng X.Anne McLean L, et al. Acid-sensing Ion Channels in Malignant Gliomas. J. Biol. Chem 2003;278:5023-5034.
35. Sandstrom PE, Jonsson O, Grankvist K, et al. Identification of potassium flux pathways and their role in the cytotoxicity of estramustine in human malignant glioma, prostatic carcinoma and pulmonary carcinoma cell lines. Eur J Cancer 1991;8:22-6.
36. Maria D, Janusz A, Pawe P, et al. Accumulation of chromosomal changes in Human glioma progression: A cytogenetic study of 50 cases. Cancer genetics and cytogenetics 1995;85:61 -67.
37. Westphal M, Hansel M, Hamel W, et al. Karyotype analyses of 20 human glioma cell lines. Acta Neurochirurgica 1994; 126:17-26.
(c) Annals of Neurosciences.All Rights Reserved