Annals of Neurosciences, Vol 12, No 2 (2005)
Annals of Neurosciences, Volume 12, Issue 2 (April), 2005
RELATIVE ROLE OF CYCLOOXYGENASE-2 (COX-2) INHIBITORS AND LIPOXYGENASE (LOX) INHIBITORS IN AGING INDUCED DEMENTIA AND OXIDATIVE DAMAGE
*Corresponding author S.K. Kulkarni, Professor of
Pharmacology Division of Pharmacology, University Institute of
Pharmaceutical Sciences, Chandigarh, India-160014
Email:
Abstract
Several inflammatory processes play a critical role in brain aging and are associated with increased vulnerability to neurodegenerati on. Cyclooxy genäse-2 (COX-2) and 5-lipoxygenase (5-LOX), two enzymes involved in the oxygenation of the arachidonic acid, are up regulated in the CNS during aging and are associated with aging related different brain pathologies. Present experiment was performed to study the effect of 5-LOX inhibitor (acetyl- keto- b- boswellic acid (AKBA), nimesulide (preferential COX-2 inhibitor) and there combination on cognitive performance of young and aged mice using plus maze test. Chronic administration of AKBA (100mg/kg p.o) and nimesulide (2.42 mg/kg,p.o.) for 15 days significantly reversed the aging induced retention deficit in mice. Co-administration of AKBA and nimesulide enhanced the cognitive performance in aged mice as compared to
Key Words: Neurodegeneration, Cyclooxygenase-2, 5-lipoxygenase, Lipid peroxidation, Gastrointestinal tolerability.
Introduction
Brain aging is associated with the increased vulnerability to the neuro-degeneration (1). A poly-unsaturated fatty acid, AA, present in the central nervous system (CNS) is main precursor for the enzymatic production of prostaglandins and leukotrienes via two pathways i.e. Cyclooxygenase (COX) pathway and lipoxygenase (LOX) pathway (2). It has been suggested that the alterations in the balance of AA and its metabolites in the CNS is associated with brain dysfunctions, including neurodegenerative pathologies of aging brain (3,4).
Reports indicate that both 5-LOX and COX-2 pathways are upregulated in the CNS and are associated with aging related brain pathologies including Alzheimer's disease (AD) (5,6). AD is a typical aging associated neurodegenerative disorder characterized by progressive deposition of β-amyloid (Aβ) in the brain resulting in the formation of neuritic plaques (7). Formation of neuritic plaques is associated with a large number of increase in immune system proteins including components of the complement cascade, cytokines and acute phase reactants, protease inhibitors and intracellular adhesion molecules (8,9). In AD neocortex displays variety of the inflammatory reaction that leads to the loss of neuronal aberration, genetic transcriptional and the cytoskeletal abnormalities, neurotransmitter deficits and characteristic lesions including neurofibrillary tangles (NFT) (10, 11). Different evidences supported the hypothesis that 5-LOX and COX-2 are the key enzymes that control the generation of pro inflammatory mediators involved in the progression of AD and drugs acting on 5-LOX and/or COX-2 may provide the pharmacological means of neuroprotection (12,13). Aging is associated with significant alterations in the pattern of DNA methylation and hormonal balance (hyperglucocorticoiduria or melatonin deficiency) that results into the up regulation of 5-LOX mRNA hence a putative involvement of 5-LOX has been hypothesized in AD (6, 14). Heterogeneity of COX-2 mRNA message abundance may reflect various degrees of expression of AD related neuro-inflammatory processes operating in both normally aging human neocortex and AD affected brain (15). Aging dependent oxidative stress is one of the mechanisms that produce vulnerability of brain to neurodegeneration. It is supposed that oxidative damage and lipid peroxidation proceeds via 5-LOX, COX and PLA2 (phospholipase A2) enzymes (16,17).
In view of the importance of 5-LOX and COX-2 enzymes as a critical neuro inflammatory mediators in the progression of AD and aging we studied the possible effectof nimesulide (preferential COX-2 inhibitor), AKBA (5-LOX inhibitor) and their combination in aging induced cognitive dysfunction and oxidative damage in mice.
Material and Methods
Animals
Young (3 months) and aged (16 months old) Swiss mice of either sex, weighing 15–20 and 35–40 g, respectively, were used. The animals were produced from Central Animals House, Panacea Biotic Ltd. Lalru, Punjab. They were housed six per cage, with free access to standard food and water, and maintained under the laboratory conditions. Mice were allowed to acclimatize in laboratory conditions before testing. Experiments were carried out between 09:00 and 18:00 h. The experimental protocols were approved by the Institutional Animal Ethical Committee and conducted according to the CPCSEA guidelines on the use and care of experimental animals.
Drugs and treatment schedule
Nimesulde, AKBA (Acetyl- keto-B-Boswellic acid) and their combination were used in the study. Nimesulide and AKBA were suspended in 0.5% carboxy methylcellulose and administered per orally (p.o.) daily for 15 days to young as well as aged mice. Control animals received only vehicle treatment (10 ml/kg, p.o.). On the day 14 after dosing, animals were tested for their sensory motor functions using an activity meter and rota-rod test. Later, their cognitive performances were assessed using elevated plus maze task. After recording the behavioral parameters mice were fasted for 24 h and their stomach was isolated and subjected for ulcerogenicity tests (pH and ulcer index).
Elevated plus maze test
The elevated plus maze was used to evaluate spatial long-term memory, following the procedure described. Briefly, the apparatus consisted of two open arms (16 × 5 cm2) and two enclosed arms (16 × 5 × 12 cm3). The arms extended from a central platform, and the maze was elevated to a height of 25 cm from the floor. On the first day, each mouse was placed at the end of an open arm. TL, the time taken by the mouse to move into one of the enclosed arms, was recorded on the first day. If the animal did not enter an enclosed arm within 90 s, it was gently pushed into one enclosed arm and the TL was assigned as 90 s. The mouse was allowed to explore the maze for 20 s, and then returned to its home cage. Retention was examined 24 h after the first day trial. Each mouse was again placed into the maze and TL was recorded. A long latency period to reach enclosed arm indicates poor retention compared to significantly shorter latencies (18).
Locomotor activity test
The animal locomotor behavioral pattern was monitored using activity meter (IMCORR India). Before subjecting the animals to cognitive tasks, they were individually placed in a Plexiglass cage (40×15×15 cm3) and the total activity count registered for 5 min. The locomotor activity was expressed in terms of total photo beams counts/5 min per animal (19).
Rota-rod test
The sensory motor performance was evaluated on a rota-rod test before subjecting the animals to cognitive tasks. The rota-rod performance was evaluated as fall off time (seconds) from the rotating rod (15 rpm/min) in a period of 5 min was recorded (19).
Studies on gastrointestinal tolerability
In order to check the chronic gastrointestinal tolerability all the animals after behavioral studies were fasted for 24 h and sacrificed, stomach isolated, gastric content collected and subjected for pH. Gastric epithelium was scored for ulcerogenicity as reported earlier (20).
Lipid peroxidation estimation (21)
Immediately after sacrificing the rats, gastric epithelium was dissected and mixed with isotonic solution. The gastric epithelium was minced properly and homogenate was prepared with 10% W/V phosphate buffer saline (0.1 M, pH=7.4) with the help of homogenizer and used to estimate the MDA levels. Lipid peroxides were measured as malonaldehyde (MDA) levels; an index of lipid peroxidation produced by acid heating using its reaction to thiobarbituric acid (TBA) was estimated. The amount of MDA formed was measured spectrophotometrically at 532 nm. The results were expressed as % lipid peroxidation compared with control taken as 100%.
Estimation of reduced glutathione levels (22)
Reduced glutathione in the gastric epithelium was estimated according to the method of Ellman. A, 0.75 ml of homogenate was precipitated with 0.75 ml of 4% sulfosalicylic acid. The samples were centrifuged at 1200-X g for 15 min at 4°C. The assay mixture contained 0.5ml of supernatant and 0.5 ml of 0.01MDTNB. The yellow color developed was read immediately at 412nm using Perkins Elmer lambda 20 spectrophotometer. The results were expressed as nmol GSH per mg protein.
Protein estimation (23)
The protein content was measured according to the method of Lowry using bovine serum albumin as standard.
Statistical analysis
Results are expressed as mean ± S.E.M. The data were analyzed using one-way ANOVA followed by Dunnett's test. The criterion for statistical significance was P <0.05.
Results
Effect of Nimesulide, AKBA and their combination on the aging-induced impairment of elevated plus mazes performance.
Control (vehicle) mice exhibited TL into the enclosed arms of 41.33±3.52s (n=6) on the first day, decreasing to 27.16±2.39 on the second day, indicating that the mice remembered the presence of the enclosed arms. In' contrast, aged mice showed no decrease in the TL on the second day, indicating that the mice did not remember the presence of the enclosed arms. Chronic treatment significantly decreased the TL on second day performance in aged mice, in comparison to young adult mice. (Fig.1).
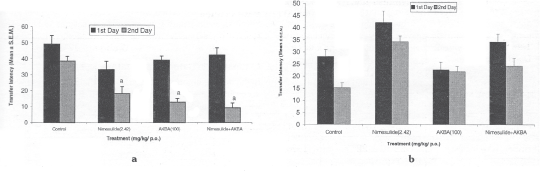
Fig. 1: Effect of chronic administration (15 days) of nimesulide (2.42mg/kg p.o.) AKBA (100 mg/kg p.o.) and their combination on transfer latency, (a) aged; (b) young mice during retention test on elevated plus maze task. aP < 0.05 compared with TL of control on 2nd day.
Chronic effect of Nimesulide, AKBA and their combination on gastrointestinal tolerability.
Chronic administration (15 days) of nimesulide (2.42 mg/kg), AKBA (100 mg/kg) and their combination showed no significant gastrotoxicity (ulcer index, pH) in comparison to control (vehicle treated) animals. (Table 1.)
| Treatment (mg/kg., p.o.) | Aged mice | Young mice | ||
|---|---|---|---|---|
| Ulcer index (Mean ± S.E.M) | pH | Ulcer index (Mean ± S.E.M) | pH | |
| Control | 0.00 ±0.0 | 4.10 | 0.00 ± 0.0 | 3.42 |
| Nimesulide (2.42) | 0.00 ±0.0 | 3.45 | 0.00 ± 0.0 | 3.84 |
| AKBA (100) | 1.00 ± 0.05 | 3.49 | 0.00 ± 0.00 | 3.62 |
| Nimesulide (2.42) + AKBA (100) | 0.00 ± 0.0 | 4.32 | 0.00 ±0.00 | 3.96 |
Effect of Nimesulide, AKBA and their combination on the locomotor activity and rota-rod performance.
In order to check the general locomotor performance of aged and adult mice before the behavioral studies, a 5 min activity test was performed for each mouse. No significant alteration in the total activity was observed in aged and young adult mice. Rota-rod performance was significantly improved in aged mice after chronic treatment with nimesulide AKBA and their combination in comparison to young mice. (Table 2 and 3).
| Treatment (mg/kg., p.o.) | Aged mice | Young mice | ||
|---|---|---|---|---|
| Basal | 15th day | Basal | 15th days | |
| Control | 183.40 ± 64.2 | 241.00 ±41.3 | 287 + 31.3 | 300 ± 0 |
| Nimesulide (2.42) | 98.54 ± 15.34 | 164.42 ± 32.3a | 246.2 ± 32.4 | 292 ± 34.3 |
| AKBA (100) | 122.60 ± 22 | 198.20 ± 34a | 300 ± 0.0 | 300 ± 0 |
| Nimesulide (2.42) + AKBA (100) |
110.24 ±30.11 | 202.42 ± 12.94a | 212.42 ± 36.21 | 300 ± 0 |
a p < 0.05 compared with control animals.
| Treatment (mg/kg., p.o.) | Aged mice | Young mice | ||
|---|---|---|---|---|
| Basal | 15th day | Basal | 15th days | |
| Control | 188.2 ± 16.62 | 160.40 ± 14.34 | 192.24 ±12.43 | 201.36 ± 8.96 |
| Nimesulide (2.42) | 176.42 ± 17.4 | 229.24 ± 18.22a | 202.24 ± 16.24 | 252.48 ± 12.84a |
| AKBA (100) | 143.2 ± 12.29 | 241.96 ± 29.26a | 213.34 ± 13.31 | 223.42 ± 14.38a |
| Nimesulide (2.42)+ AKBA (100) | 138.46 ± 13.46 | 198.96 ± 18.14a | 182.26 ± 8.94 | 226.42 ± 18.24a |
a p < 0.05 compared with control animals.
Lipid peroxidation
Aged mice showed significant increase in lipid peroxidation in comparison to young mice. Nimesulide (2.42 mg/kg) and AKBA (100 mg/kg) significantly decreased the lipid peroxidation. Further, their combination (nimesulide 2.42 mg/kg) + AKBA (100 mg/kg) showed more pronounced effect when compared with their per se treatment (Fig. 2).
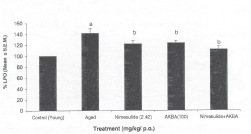
Fig. 2: Effect of chronic administration (15 days) of nimesulide (2.42mg/kg p.o.) AKBA (100 mg/kg p.o.) and their combination on increased lipid peroxidation in aged mice. * P < 0.05 compared with control young mice #P < 0.05 compared with control aged mice.
Reduced glutathione levels
Aged mice showed significant decrease in reduced glutathione level in comparison to young mice. Nimesulide (2.42 mg/kg) and AKBA (100 mgfcg) significantly decreased the reduced glutathione level. Further, their combination (nimesulide 2.42 mg/kg) + AKBA (100 mg/kg) showed more pronounced effect when compared with their per se treatment (Fig. 3).
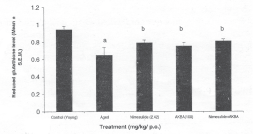
Fig. 3: Effect of chronic administration (15 days) of nimesulide (2.42mg/kg p.o.) AKBA (100 mg/kg p.o.) and their combination on decreased reduced glutathione level in aged mice., * P < 0.05 compared with control young mice #P < 0.05 compared with control aged mice.
Discussion
Cognitive impairment such as learning deficits and delayed amnesia are the most happening age related changes in human beings and animals. In search for the pathogenic mechanisms of AD, much interest has been recently focused on brain inflammation. The appearance of activated microglia, reactive astrocytes component protein and proinflammatory cytokines associated with neurofibrillary tangles and β-myeloid in AD affected neocortex suggests that AD affected brain may be in chronic state of inflammation. Recently, it is reported that two of the inflammatory isoenzymes COX-2 and 5-LOX (mRNAs and proteins) are increased in the brain of the aging animals and suggest a role of these enzymes in neurodegeneration (6).
COX activity was found to be greater in the older rats (24). It has been suggested that increased COX activity with age is due to the activation of COX catalytic reactions, presumably by reactive oxygen species (24). Neuronal expression of COX-2 is increased in course of seizures, stroke and neuroprotection was demonstrated by COX inhibitors in several models of neurodegeneration both invitroand invivo(6,25). Further support for the participation of COX-2 in these disorders was provided by experiments in transgenic mice. COX-2 knock out mice exhibited a reduced severity in different models of neurodegeneration (26), where as those over expressing COX-2 in neurons showed age-dependent cognitive deficits, neuronal apoptosis suggesting the role of neuronal COX-2 in pathophysiology of these disorders (27).
5-LOX expression is also greater in older rats than the younger rats (28). 5-LOX is one of the predominant up regulated gene during hippocampal excitotoxicity, multiple sclerosis, reperfusion of the ischaemic brain (stroke), and transmissible spongiform encephalopathies (mad cow disease). It was found that 5-LOX pathway is crucially involved in these disorders and 5-LOX inhibition could act as neuroprotectives (29,30).
Enzymatic activity is not the only mechanism by which COX-2 and 5-LOX can influence the normal brain physiology. 5-LOX plays a regulatory role in the binding of coactosin like protein (CLP) to F-actin (31,32) and possibly also in the F-actin stability. F-actin stability in dendritic spines of CNS neurons maintains the stability of synaptic structures and is disrupted by neurodegeneration. This, interplay between 5-LOX, CLP and F-actin might point to a novel mechanism of neuronal degeneration (6,14,34). Certain COX inhibitors are also reported to Set via mechanism unrelated to COX-inhibition. Indomethacin, ibuprofen and Sulindac sulphate reduce Aβ42 peptide production in a variety of cells and lowered the mice brain levels of Aβ42.
Based on these, present study demonstrates the chronic administration of nimesulide (2.42mg/kg p.o.) a preferential COX-2 inhibitor and AKBA (100mg/kg p.o.) a 5-LOX inhibitor to aged mice for 15 days improved the cognitive performance in one trial spatial and passive avoidance tasks significantly as compared to untreated aged animals. Further the co-administration of nimesulide and AKBA significantly potentiates the cognitive performance on elevated plus maze and step through paradigm where as they did not improve the cognitive functions in young animals. Results from this investigation is in accordance to the previous reports that explain the role of both enzymes in aging related pathologies and suggested that inhibition of COX-2 and/ or 5-LOX could be a potential neuroprotective therapy.
Neurodegenerative diseases involve the neuronal death. Oxidative stress induces age dependent neuronal apoptosis, which is one of the mechanism that produce age dependent vulnerability to neurodegeneration. Oxidative stress also favours lipid peroxidation in biomembrahes, which proceeds enzymatically via PLA2, 5-LOX and COX-2 (16,17). Considering the fact that increased oxidative stress and lipid peroxidation is associated with increased COX-2 and 5-LOX metabolites we studied the effect of Nimesulide, (2.42mg/kg p.o.) and AKBA (100mg/kg p.o.) and their combination on aging related increase in lipid peroxidation and oxidative stress. Aging resulted into significant increase in lipid peroxidation and reduction in reduced glutathione levels when compared to young mice. Chronic treatment with nimesulide (2.42mg/kg p.o.) and AKBA (100mg/kg p.o.) prevented the oxidative stress and lipid peroxidation significantly. Further co administration of AKBA and nimesulide significantly increased the effect as compared to
Taken together these data are indicative to the fact that aging is associated with increased memory deficits and oxidative stress which could be because of over expression of COX-2 and/or 5-LOX isoenzymes. These effects could be reversed by the chronic treatment with COX-2 inhibitor or 5-LOX inhibitor by virtue of their enzymatic or non-enzymatic action. Further the combined inhibition of both the isoenzymes will significantly increase the protective effect as compared to their per se effect.
In conclusion, the study provides an evidence for the development of newer protective therapies targeting COX-2,5-LOX or both of them (dual inhibitors) for the treatment of aging associated brain disorders such as Alzheimer's diseases.
References
1. Manev H, Uz T Lipoxygenase, Cyclooxygenases, and the Aging Brain Comments of Theoretical biology 2001; 6:447–459.
2. Funk CD. Prostaglandins and leutrotrienes: advances in eicosanoid biology. Science 2001; 294: 1871–1875.
3. Pect M, Glen I, Horrobin DE Phospholipid spectrum disorder in psychiatry. Lancashire: Marius press 1999.
4. Pect M, Glen I, Horrobin DF. Phospholipid spectrum disorder in psychiatry. Lancashire: Marius press 1999.
5. Uz T, Pesold C, Longone P, Manev H. Aging-associated up regulation of neuronal 5-lipoxygenass expression: Putative role in neuronal vulnerability. FASEB J 1998; 12: 439–449.
6. Manev H, Uz T, Sugaya K, Qu T. Putative fole of neuronal 5-lipoxygenase in an aging brain. FASEB J. 2000; 14: 1464–1469.
7. Geula C, Saroff D, Lorenzo A, Yuan M, Yankner BA Aging renders the brain vulnerable to amyloid ?-protein neurotoxicity. Nature Med 1998; 4: 827–831.
8. Rogers J, Styren SD, Civin WH. Expression of immune system associated antigens by cells of the human central nervous system: relationship to pathology of Alzheimer's diseases. Neurobiol. Aging 1998; 9: 339–349.
9. Mcgeer PL, Mcgeer EG. The inflammatory response system of brain: implications for therapy in Alzheimers and other neurodegenerative diseases. Brain Res Rev.1995; 21: 195–218.
10. Blain H, Jouzeau JY, Blain A, Terlain B, Trechot P, Touchon J, Jeandel C. Alzheimer's diseases and nonsteroidal anti inflammatory drugs with selectivity of cyclooxygenase-2: rationale and perspectives. Presse Medicale 2000; 29: 267–273.
11. Combs CK, Johnson DE, Karlo JC, Cannady SB. Inflammatory mechanisms in Alzheimer's diseases: inhibition of ?-amyloid stimulated pro inflammatory responses and neurotoxicity by PRAR gamma agonists. J Neurosci 2000; 20: 558–567.
12. Sugaya K, Uz T, Kumar V, Manev H. New anti-inflammatory treatment strategy in Alzheimer's disease. Jpn J Pharmacol.2000; 82: 85–94.
13. Baran H, Vass K, Lassmann H, Hornykiewicz O. The cycloxygenase and lipoxygenase inhibitor BW755C protects rats against kainioacid – induced seizures and neurotoxicity. Brain Research 1994; 646: 201–206.
14. Qu T, Manev R, Manev H. 5-lipoxygenase promotor polymorphism in patients with early-onset and late-onset Alzheimer's disease. J Neuropsychiatry clin Neurosa-2001; 13: 304–305.
15. Lukiw WJ, Bazan NG. Cyclooxygenase-2 (COX-2) mRNA abundance, stability and hyper activity in Sporadic Alzheimer's neocortex. J Neurol Res 1997; 50: 937–945.
16. Kim GW, Chan PH. Oxidative Stress and neuronal DNA fragmentation mediate age-dependent vulnerability to the mitochondrial toxin, 3-nitropropionic acids, in the mouse straitum. Neurobiol Dis 2001; 8: 114–126.
17. Farooqui AA, Ong WY, Lu HR, Horrocks L.A. Neurochemical consequences of kainate induced toxicity in brain: Involvement of arachidonic acid release and prevention to toxicity by phospholipase A2 inhibitors. Brain Res Rev 2001; 38: 61–78.
18. Itoh J, Nabeshima T, Kameyama T. Utility of an elevated plusmaze for dissociationof amnesic and behavioural effects of drugs in mice. Eur J Pharmacol 1991; 1994: 71–76.
19. Reddy DS, Kulkarni SK. Possible role of nitric oxide in the nootropic and antiamnesic effects of neurosteroids on aging and dizocilpine induced learning impairment. Brain Res 1998; 799: 215–219.
20. Jain NK, Singh A, Kulkarni SK. Analgesic, anti inflammatory and ulcerogenic activity of a zinc-naproxen complex in mice and rats. Pharm Pharmacol Communl999; 5: 599–602.
21. Wills ED. Mechanism of lipid peroxide formation in animal tissues. Biochem Jour 1965; 99: 667–676.
22. Ellman GL. Tissue sulfhydryl groups. Arch of Biochem Biophy.1959; 82: 70–77.
23. Lowry OH, Rosenbrought NJ, Farr AL, Randall RJ. Protein measurements with the Folin-phenol reagent. J Biol Chem 1951; 193: 265–275.
24. Beak BS, Kim JW, Lee JH, Know HJ, Kim ND, Kang HS, Chung HY. Age related increase of brain cyclooxygenase activity and dietary modulation of oxidative stress J Gerontol 2001; 56 : B426-B431.
25. Hoffmann C. COX-2 in brain and spinal cord-Implications for therapeutic use. Curr Med Chem 2000; 7: 113–120.
26. Iadecola C, Niwa K, Araki E, Ross ME. Reduced susceptibility to ischemic brain injury and NMDA mediated neurotoxicity in cyclooxygenase deficient mice. Proc Natl Acad Sci USA. 2001; 1294–1299.
27. Andreasson K, Zhang Y, Shaffer A, Isakson R Age dependent cognitive deficits and neuronal apoptosis in cyclooxygenase-2 transgenic mide. J. Neurosci.2001; 21: 8198–8209.
28. Qu T, Uz T, Manev H. Inflammatory 5-LOX mRNA and protein are increased in brain of aging rats. Neurobiol. Aging 2000; 21: 647–652.
29. Shishido Y, Furushiro M, Hashimoto S, Yokokura T. Effect of nordihydroguaiareticacid on behavioural impairment and neuronal cell death after forebrain ischemia.Pharmacol Biochem Behav 2001; 69: 469–474.
30. Stewart LR, White AR, Jobling MF, Maher F, Collins SJ, Cappai R. Involvement of the 5-lipoxygenase pathway in the neurotoxicity of the prion peptide Pr P 106–126. J Neurosci Res 2001; 65: 565–572.
31. Radmark O, Samuelsson B, Provost R Interaction of 5-lipoxygenase with cellulsar proteins. Proc Natl Acad Sci USA. 1999; 96: 1881–1885.
32. Provost P, Doucet J, Samuelsson B, Radmark O. 5-lipoxygenase interacts with coactosin-like proteins. J Biol Chem 2001; 276: 16520–16527.
33. Zhang W, Han SW, McKeel DW, Wu JY Interaction of presenilis with the filamin family of actin-binding proteins. J Neurosci 1998; 18: 914–922.
(c) Annals of Neurosciences.All Rights Reserved